| Главная | Библиотека | Форум | Гостевая книга |
ИНТЕЛЛЕКТУАЛЬНЫЕ
СПОСОБНОСТИ
И ГРУППОВОЕ
ПОВЕДЕНИЕ

Муравей обладает не только ощущениями, но и рассудком, разумом, памятью. Цицерон. О природе богов Я отметил, сколь счастливо, кажется, живут вместе члены одного сообщества. Высшая гармония царит повсюду. Маленький народец даже помогает друг другу при нужде или затруднении. Когда один голоден, другой кормит его. Когда один нездоров, другой помогает ему. Рабочих поменьше... переносят, нежно захватывая, их более выносливые собратья. У. Уайт. Муравьи и их обычаи |
ГРАНИЦЫ ИНТЕЛЛЕКТА
Человекообразные обезьяны, без сомнения, больше других животных приближаются к человеку по строению тела, но, рассматривая жизнь муравьев, их общественную организацию, их обширные сообщества, тщательно устроенные жилища и дороги, их владение домашними животными, а в некоторых случаях даже и рабами, нельзя не признать, что по своей разумности они имеют полное право занимать место сразу же после человека. Дж. Лаббок. Муравьи, пчелы и осы Иначе у муравьев. У них нет ни разума, ни самосознания, и поэтому они вовсе не способны копаться в темных вопросах «откуда» и «зачем». Э. Васманн. Итоги сравнительной психологии |
Есть ли разум у насекомых? В течение многих столетий большинство ученых
отвечало на этот вопрос положительно. В науке господствовал антропоморфизм
— наделение
животных человеческими свойствами. Истоком этого было обыденное сознание
людей, объяснявших поступки животных, исходя из собственных мотивов и
побуждений. Казалось очевидным, что насекомое может радоваться и огорчаться,
любить
и испытывать неприязнь, рассчитывать свои действия и строить планы, приобретать
опыт и делать выводы. Тем более общественные насекомые — пчелы, термиты,
муравьи — с их удивительно сложными жилищами, разведением грибов и домашнего
скота и даже рабовладением. Конечно же, их поведение разумно! Примером
такого
натурфилософского подхода может служить первый эпиграф.
Но уже в XVI в. возникла и прямо противоположная точка зрения, впервые
высказанная испанским врачом Г. Перейрой и получившая развитие в трудах
известного философа
Р. Декарта. Согласно этому взгляду, животное — машина, действующая чисто
автоматически. Французский натуралист Ж. Бюффон в своей 36-томной «Естественной
истории» (1749—1804) описывал муравьев, пчел и других общественных насекомых
как механизмы, неспособные мыслить и анализировать, однотипно реагирующие
на внешние стимулы. На рубеже XIX и XX вв. известный физиолог и исследователь
поведения А. Бете создал на основе этих взглядов детальную теорию поведения
общест-
351
венных насекомых, в которой рассматривал их как «рефлекторные машины»,
автоматы, действующие, повинуясь только внешним стимулам, реагируя
на них строго определенным
образом.
Две эти крайние точки зрения долго существовали в науке, поскольку
обе стороны ограничивались общими рассуждениями и не пытались опытными
данными
обосновать
их. Только со второй половины XIX в. началась эра экспериментального
изучения поведения животных. В 80-х годах в научной среде развернулась
бурная дискуссия
о наличии разума у общественных насекомых. Ее участники — Э. Васманн,
А. Бете, Дж. Лаббок, О. Форель, К. Эмери и др. — приводили все новые
и новые
полученные ими фактические данные. Выяснилось, что муравьи и пчелы,
как и высшие позвоночные, способны быстро обучаться и использовать
приобретенные
навыки в своей жизни. Но в то же время оказалось, что поведение насекомых
в значительной степени определяется инстинктами, т. е. является врожденным.
Было установлено, что общественные насекомые способны передавать
друг другу некоторые сообщения, но не могут использовать свой «язык»
как
средство мышления,
создавать понятия и представления.
Что же касается разумности, то одни ученые (Бете, Васманн) продолжали
считать ее прерогативой человека, имеющей божественное происхождение,
другие же
(Форель, Эмери), придерживаясь позиций дарвинизма, утверждали, что
у высших, животных,
в том числе и у муравьев, являющихся вершиной эволюции насекомых,
есть некоторые элементы разумной деятельности. Но убедительных доказательств
своих позиций
обе стороны привести не могли. Дискуссия в основном завершилась к
началу
XX в., хотя многие разногласия существуют в науке до сих пор.
Одно из расхождений касалось соотношения между врожденными (инстинктивными)
и приобретенными формами поведения у насекомых. А. Бете
и Ж. Леб, например, утверждали, что в жизни насекомых все определяется
инстинктами
и роль
обучения незначительна, а выдающийся американский психолог и энтомолог
Т. Шнейрла
доказывал, что инстинктов в чистом виде не существует и все формы
поведения насекомых создаются под воздействием внешней среды в течение
жизни.
Теперь этот спор представляет чисто исторический интерес. Развитие
генетики
показало, что нельзя противопоставлять врожденные и приобретенные
формы поведения.
Все признаки и свойства организма, в том числе и специфические особенности
его поведения, имеют генетическую основу, но появляются и формируются
у развивающегося организма при обязательном участии окружающих условий.
Нельзя
найти такую
форму поведения, на которую генетические задатки, т. е. генотип животного,
или, напротив, внешняя среда не оказывали бы никакого влияния.
352
Можно говорить только о степени обусловленности той или иной формы
поведения генотипом и средой. Рассмотрим с этой точки зрения
поведение общественных
насекомых.
Прежде всего надо сказать, что поведение общественных насекомых,
в том числе и самых высокоорганизованных, в значительно большей
степени определяется
наследственными факторами, чем влиянием внешней среды. Многие
формы социального поведения не требуют для своего развития какого-либо
обучения,
передачи
навыков
от сородичей. Возьмем, например, соты медоносной пчелы с запечатанными
куколками и поставим в пустой улей. Вскоре в нем появятся молодые
пчелы. Они начнут
чистить ячейки, строить новые соты, заботиться о личинках (если
мы дадим им соты с яйцами), вылетать за кормом, хотя никогда
не
встречались
со
старыми и опытными особями, от которых могли бы научиться делать
все это. Увидим
мы в экспериментальном улье и танцы, ими пчелы будут сообщать
друг другу о найденном корме. Значит, способность пчел передавать
и
понимать сигналы
тоже инстинктивна и не приобретается в течение жизни от сородичей.
Ведь нельзя же серьезно говорить о том, что пчелы учатся танцевать,
сигнализировать
об
опасности, просить и предлагать корм, будучи еще личинками. Какой
контраст с поведением человека! Мы с вами получаем практически
все социальные
навыки поведения и общения путем обучения и подражания другим
людям. Язык человека
развивается тоже только при общении со взрослыми с первых лет
жизни (вспомните не выдуманного, а настоящих маугли — детей,
выросших
в полной изоляции
и потому по уровню психического развития стоящих ниже пещерного
человека!).
Эксперименты, подобные описанному, хорошо удаются и с пчелами
других видов, и с муравьями. Иногда их проделывает сама природа.
Например,
муравьи-рабы, попавшие в гнездо рабовладельцев на стадии личинки
или куколки и не общавшиеся
со взрослыми насекомыми своего вида, проявляют видоспецифичное
поведение во всех отношениях. Поэтому гнезда муравьев-амазонок
находящиеся
в рабстве бурый лесной, прыткий или песчаный муравьи строят по-разному,
но в строгом
соответствии со стандартами своего вида.
Когда в гнездо одного из видов безжалых пчел поместили несколько
сотен коконов другого вида, то вскоре возникла колония со смешанным
населением
рабочих.
И оказалось, что подсаженные на стадии куколок пчелы строят ячейки
для провизии и соты, типичные по форме и размерам для своего
вида. В результате
получилось
«гибридное» по структуре гнездо. Значит, строительное поведение
тоже врожденное, ведь не учатся же пчелы строить на стадии личинки!
В
этом гнезде обнаружилось
еще одно интересное явление. Пчелы, развивающиеся в ячейках большего
размера, сооруженных рабочими другого вида, были крупнее, но
сами возводили ячейки,
типичные для
353
своего вида и по размеру, и по форме. Вот насколько устойчив
врожденный видовой стереотип строительного поведения!
Еще в одном эксперименте сот с яйцами, отложенными маткой
краинской расы медоносной пчелы, поставили в улей итальянской
расы. Светлоокрашенные
итальянки выкормили гораздо более темных краинских пчел —
возникла смешанная
семья.
Тогда исследователи стали наблюдать за танцами, строго фиксируя
их параметры. И выяснилось, что краинские пчелы танцевали
в полном соответствии
с «диалектом»
своей расы. А надо сказать, что краинская раса — наиболее
живо танцующая среди всех исследованных рас: при одном и
том же
расстоянии до кормушки
ее разведчицы совершают виляющий пробег на 15—20% быстрее,
чем пчелы-итальянки. Звуковые сигналы дальности, издаваемые
во время
танца, у выкормленных
итальянками краинских пчел тоже ничем не отличались от типичных
для этой расы. Значит,
основные параметры танца определяются наследственностью и
не изменяются при
выращивании личинок пчелами другой расы.
Ну, а как обстоит дело с восприятием информации, заключенной
в танце? Краинским пчелам из смешанной семьи предложили кормушку,
удаленную
на 500 м от улья.
Их танцы совершенно правильно поняли другие краинские пчелы,
вскоре
появившиеся на той же кормушке. Но вот итальянки ошиблись,
и почти все прилетели
на пустую контрольную кормушку, находившуюся в 300 м от улья.
Дело в том,
что на их
«диалекте» темп, в котором танцевали краинские пчелы, указывает
расстояние в 300, а не 500 м. Когда же танцевали итальянки,
то просчет допускали
краинские пчелы, но уже в большую сторону! Значит, и способность
«расшифровывать» танцы врожденная и не зависит от воспитания
и обучения.
Преобладание у общественных насекомых врожденных: элементов
поведения, повторим, не свидетельствует, что внешняя среда
и опыт не играют
в их жизни никакой
роли. Напротив, они оказались очень способными учениками,
нисколько не хуже, чем птицы и млекопитающие. Особенно подробно
исследовано
обучение у медоносной
пчелы. Условные рефлексы образуются у нее нередко сразу,
т. е. при единственном
сочетании безусловного стимула (пищи) с условным, в качестве
которого могут выступать цвет, форма и расположение предметов,
запахи, звуки,
осязательные
сигналы и даже время. Да, да, пчелы легко обучаются прилетать
на кормушку в определенное время суток и помнят об этом в
течение многих дней.
Пчеле достаточно всего три секунды пососать сахарный сироп,
чтобы запомнить цвет или запах кормушки, на которой она получила
пищевое
подкрепление.
Ничуть
не хуже образуются условные рефлексы у муравьев, ос и шмелей.
Очень хорошо обучаются муравьи, пчелы и шмели находить правильный
путь в запутанных лабиринтах, причем иногда му-
354
равьи решают эту задачу столь же быстро, как и крысы, славящиеся
своей сообразительностью. У муравьев удается выработать
даже довольно сложные
инструментальные рефлексы.
В эксперименте А. Рехштайнера возле дороги малого лесного
муравья был установлен аппарат, две нити которого соединялись
со шторками.
Если
муравьи тянули
первую нить, то получали в качестве вознаграждения сахарный
сироп за шторкой; потягивание
второй нити не поощрялось. Муравьи скоро уловили эту
закономерность и дергали за первую нить свыше 60 раз в час, а за вторую
— не более 20.
Причем нередко
за нить тянули одни муравьи, а сахарный сироп поедали
другие.
Высокоорганизованные общественные насекомые значительно,
быстрее обучаются, чем многие одиночные. Недаром у муравьев
и пчел
заметно лучше по сравнению
с близкородственными, но ведущими одиночный образ жизни
насекомыми развиты так называемые грибовидные тела —
отделы мозга, отвечающие
за высшую
нервную деятельность, в том числе и за обучение. У медоносной
пчелы грибовидные тела составляют примерно 1/6 объема
мозга, а у муравьев
рода формика
—
1/3 часть.
Да и сам головной мозг занимает у общественных видов
гораздо большую часть от объема тела, чем у одиночных
насекомых.
Во всех экспериментах по образованию условных рефлексов
и обучению в лабиринте оказывается, что насекомые вовсе
не
одинаковы по
своим способностям:
среди
них есть очень сообразительные и довольно тупые, но основную
массу составляют середнячки. Этот факт заслуживает особого
внимания, поскольку подтверждает
представления о неравноценности психических качеств и
склонностей насекомых, которые мы уже обсуждали в связи
с вопросом
о разделении труда.
Как же используют общественные насекомые свои способности
к обучению в повседневной жизни? Прежде всего, конечно,
при ориентации
на
кормовом участке
и добывании
пищи. Пчелы улетают от гнезда на многие километры, осы
и шмели — на сотни метров. Муравьи уходят не так далеко,
но
передвигаются
пешком,
что несравненно
сложнее. Для того чтобы вернуться в гнездо, нужно обладать
отличной топографической памятью, знать множество ориентиров
на кормовом
участке и вокруг гнезда.
Если улей с пчелами ночью перевезти на новое место, то
утром его обитатели,
вылетая за пищей, совершат несколько все расширяющихся
кругов над своим жилищем и отлично запомнят все ориентиры,
необходимые
им
для возвращения.
Феноменальные
способности! Очень многие люди в такой ситуации непременно
заблудились бы.
Обнаружив солидный источник корма, пчела или муравей
прочно удерживают в памяти это место и способны вновь
найти его.
Нередко насекомое
«берет на
заметку» множество таких «перспективных участков» и периодически
посещает их в надежде на появление там пищи. Способность
к образованию
355
условных рефлексов на время позволяет пчелам прилетать
к тем или иным медоносам как раз тогда, когда они
начинают выделять
нектар
(а у разных
растений
это происходит в разную пору суток).
Поведение насекомых при поисках и добывании корма
в основном является врожденным. Но приспособление
к конкретным
условиям
местности,
определенной пище или
виду добычи, оказывается, невозможно без накопления
индивидуального опыта. Теперь известно, что шмели
и пчелы-новички, впервые
вылетающие за кормом,
ведут себя на цветках неуверенно, возятся дольше,
чем опытные сборщицы, и работают, так сказать, с
меньшей
производительностью
труда. Ловкость
они
приобретают довольно быстро. Но и искушенные пчелы,
попадая на еще незнакомый им вид растения, затрачивают
некоторое
время на
знакомство
с устройством
его цветка и освоение эффективных приемов сбора пыльцы
и нектара. Как мы уже знаем, общественные пчелы в
отличие от
большинства
одиночных насекомых не имеют узкого круга кормовых
растений и берут взяток
практически
со
всех
медоносов в данной местности. Цветки растений устроены
настолько разнообразно, что невозможно обладать правилами
поведения,
пригодными для всех видов.
Поэтому пчелы от рождения владеют только основными
приемами сбора нектара и пыльцы
и приспосабливают их к конкретным цветкам путем тренировки
и накопления опыта.
Некоторые наблюдения позволяют предполагать, что
и для муравьев, по крайней мере отдельных видов,
индивидуальный
опыт может
играть существенную
роль
в «овладении профессией» охотника, сборщика семян
или пади, строителя, водоноса или няньки. К сожалению,
пока
мы практически
ничего
не знаем об этой стороне
жизни общественных насекомых. Такие вещи гораздо
легче
принимать на веру или отрицать, чем доказать их существование
или отсутствие.
Лишь
в последние
годы стали появляться первые работы в этом направлении.
Я. Добржанская, наблюдая за поведением муравьев-амазонок
и их рабов, выяснила, что при взаимодействии между
ними важное значение
имеет
приобретение опыта,
приспособление друг к другу. В начале лета, при первых
набегах за куколками, поведение амазонок и рабов
еще не скоординировано.
Это
и понятно, ведь
и среди рабов, и среди хозяев больше всего рабочих,
появившихся из куколок прошлым летом и потому неопытных.
Выход колонны
амазонок влечет за собой
сильное возбуждение рабов, которые бестолково мечутся
вокруг муравейника и становятся крайне агрессивными.
Когда рабовладельцы
возвращаются
с добычей, возле гнезда возникает жуткая давка —
многочисленные амазонки
с куколками
не могут сразу все войти внутрь через узкие отверстия.
Рабы — бурые лесные
муравьи — бросаются срочно расширять входы, чтобы
постепенно пропустить в
гнездо все полчище амазонок. С каждым новым
356
походом рабы приступают к строительным работам все
раньше и раньше и примерно после десятого набега
начинают расширять
входы сразу
же, как
только амазонки
уйдут за куколками. Они уже не носятся беспорядочно
около гнезда,
проявляя бессмысленную агрессивность, а занимаются
нужным делом. И когда армия
рабовладельцев возвращается, все уже подготовлено
к встрече и амазонки быстро и без сутолоки
входят в гнездо.
Несколько иначе складываются взаимоотношения
амазонок с песчаными муравьями. В этом случае
рабовладельцы
поначалу бросают доставленных
ими куколок
на муравейник, а рабы подбирают их и уносят внутрь.
Отдельные рабы пытаются взять коконы непосредственно
у хозяев,
но
те не отдают
свою ношу; иногда
рабы даже втаскивают в гнездо вместе с куколкой
упирающуюся амазонку. С
каждым набегом взаимодействия песчаных муравьев
с хозяевами все более отлаживаются. И вот примерно
после десятого
похода почти
все амазонки
сразу передают
куколок
рабам «из рук в руки», а те уносят их в гнездо.
Ж. И. Резникова и Л. Л. Васильева изучали процесс
«взросления» вышедших из куколок рабочих прыткого
муравья и кровавого
муравья-рабовладельца. Первый
вид характеризуется несложным социальным поведением,
простыми формами коммуникации, фуражиры его в
основном охотятся
в одиночку. Социальное
поведение рабовладельца
гораздо более развито: муравьи совершают организованные
набеги за
рабами, коллективно добывают корм, владеют разнообразными
формами сигнализации.
Оказалось, что прыткие муравьи почти сразу после
освобождения из коконов начинают ухаживать
за расплодом и уже на третий день по поведению
практически не отличаются от взрослых рабочих.
Рабовладельцы
взрослеют медленнее.
В первые
дни после появления из коконов они беспомощны
и нуждаются в уходе более
взрослых собратьев. Вначале они даже неспособны
координировать движения усиков
и
полностью овладевают
ими только к шестому дню. Примерно с этого же
срока молодые муравьи начинают самостоятельно
питаться
сахарным сиропом.
Известно,
что чем сложнее социальное
поведение животных, тем более растянуто у них
детство и юность, во время которых потомство
приобретает
индивидуальный опыт.
Как это
ни удивительно,
но и у муравьев действуют те же закономерности.
Чемпионом же по продолжительности периода взросления
является,
конечно,
человек.
Дальнейшие эксперименты продемонстрировали, как
муравьи приобретают опыт в процессе взросления.
Выяснилось,
что основные поведенческие
реакции
быстрее всего формируются у муравьев, живущих
с момента выхода из коконов в материнском
гнезде, несколько медленнее — у муравьев, «воспитывавшихся»
в отдельных садках взрослыми рабочими, и еще
медленнее — у муравьев,
изолированных
от старых
рабочих с самого начала. В последнем случае молодые
муравьи длительное время
357
неспособны даже к трофаллаксису и не реагируют
на соответствующие сигналы собратьев. Но
когда молодых
муравьев, содержавшихся
в изоляции группами
по 10 особей, регулярно «выгуливали», т.
е. давали им возможность побродить по открытой площадке,
встречаясь с разнообразными
объектами, взросление
их
значительно убыстрялось по сравнению с лишенными
этого моциона муравьями. Таким образом, все
основные формы
поведения у
муравьев являются врожденными,
не требуют специального обучения и определяются
процессом созревания. Но созревание намного
ускоряется, если
молодые
муравьи живут
вместе со взрослыми
или имеют более широкие возможности для приобретения
индивидуального опыта.
Роль опыта в формировании социального поведения
подчеркивают и результаты экспериментов по
обучению в раннем возрасте
— импринтингу. Мы уже
говорили о том, что муравьи запоминают запах
окружающих их рабочих и царицы
в первые дни после выхода из куколок, а может
быть, даже и на стадии
личинки.
Оказывается,
и запах расплода своего вида они узнают именно
в это время. Когда вышедших из куколок обыкновенных
лесных
муравьев
итальянский энтомолог Ф. Ле
Моли додержал первые две недели вместе с
куколками своего вида, то они в дальнейшем
хорошо их узнавали и ухаживали за ними, а
куколок
других видов поедали. Если же они проводили
это время
вместе
с куколками волосистого
лесного
муравья,
то потом считали «своим» его расплод, а куколок
собственного вида пожирали. Такие же результаты
были получены
и в опытах с мирмиками.
Выяснилось даже, что импринтинг не только
влияет на способность узнавать расплод, но
и участвует
в формировании
стремления
заботиться о потомстве.
Если рабочих обыкновенного лесного муравья
первые три недели их жизни содержать без
расплода, то
в дальнейшем
они не
то что не
будут отдавать
предпочтение
своим или чужим куколкам, но и вообще перестанут
заботиться о них, бросив на произвол судьбы.
И вот что интересно.
Если к таким
муравьям
подсадить
опытных рабочих, которые унесут в гнездо
валяющиеся на арене коконы и начнут ухаживать
за ними,
то нерадивые няньки через
несколько
дней «одумаются»
и постепенно будут оказывать коконам все
больше внимания. В конце концов врожденные
элементы заботы о потомстве у них полностью
восстановятся. Не является ли это приобретением
опыта путем
подражания? Или активная
деятельность
взрослых
рабочих просто пробуждает врожденные способности
молодых?
Импринтинг, вероятно, широко распространен
у общественных насекомых. Если рабочие рыжей
мирмики
в первые
дни после выхода из куколок
встречаются с царицами, то позже, будучи
няньками, они станут активно препятствовать
развитию
личинок в крылатых самок в присутствии царицы.
Но если му-
358
равьи «повзрослеют», не ведая о существовании
цариц, то в дальнейшем будут гораздо
слабее реагировать на них в
гнезде
и изменять
в соответствии с
этим свое отношение к личинкам. Значит,
у мирмик происходит импринтинг царицы.
Молодые самки муравьев-амазонок, основывая
новые гнезда путем временного социального
паразитизма, предпочитают
проникать
в гнезда того вида,
который содержался в их родном гнезде
в качестве рабов. Наверное, они запоминают
запах рабов и затем ориентируются на
него в своих
поисках.
Мы говорили уже о том, что коммуникация
посредством танцев у медоносных пчел
является врожденным
поведением. Но
все-таки обучение
играет
и здесь некоторую
роль. Эксперименты ленинградской исследовательницы
Н. Г. Лопатиной позволили установить,
что молодые пчелы, начиная
танцевать,
немного занижают расстояние,
а параметры их танца довольно изменчивы.
Однако по мере
того, как они осваивают путь к кормушке,
танец их приобретает все
большую устойчивость и указание
расстояния становится правильнее. В дальнейшем
опытные пчелы танцуют
стабильно, точно сообщая о дистанции.
Следовательно, пчеле необходимо некоторое
время,
чтобы научиться безошибочно соотносить
темп своего танца с путем, который
она пролетает от кормушки к улью. Сама
же по себе способность танцевать, изменять
темп
и
ориентацию танца в соответствии
с расстоянием до
корма и направлением полета к нему является
врожденной.
Восприятие информации, сообщаемой в танцах,
тоже требует опыта. Оказалось, что пчелы,
которые в течение своей
жизни не встречались
с танцующими
разведчицами, почти индифферентны и к
настоящим танцам, и к модели танцующей
пчелы. Они
не расценивают танец как сигнал. Нужно
время, чтобы пчела уловила связь между
танцами и
наличием корма
и стала
проявлять специфическое
внимание
к танцующим
пчелам. Например, ко многим запахам молодые
пчелы
относятся нейтрально. Но стоит пчеле
хотя бы один раз получить
от танцующей разведчицы
или просто сборщицы капельку нектара
с определенным запахом, и она будет
отныне воспринимать
этот запах как сигнал о наличии пищи.
Так же она будет относиться к этому запаху
и на теле
танцующих
пчел.
Конечно, способности
пчел следовать
за танцовщицами,
оценивать темп и другие параметры их
танцев
и руководствоваться этими указаниями
в полете к
источнику корма являются
врожденными. Но для
того,
чтобы эти
способности обнаружились и были правильно
использованы, необходимо обучение, приобретение
опыта.
Еще более интересные результаты были
получены Г. М. Длусским при изучении
значения предшествующего
опыта
в мобилизации
на пищу у
рыжей мирмики.
Экспериментальные группы составляли молодые,
еще
не занимавшиеся фуражировкой рабочие,
а также царицы и расплод из одной крупной
природ-
359
ной колонии мирмики. Группы получали
равное количество корма, но были
поставлены в
разные условия фуражировки.
В первой
(контрольной) группе
пищу муравьям
давали в кормушках на неограниченное
время. Во второй группе кормление
проводили так же, но в полной темноте.
В третьем варианте кормушку выставляли
только
на один час каждый день и муравьям
приходилось поторапливаться, чтобы успеть обеспечить
себя
и расплод пищей. И,
наконец, в четвертой и
пятой группах
животную пищу (куколки лазиусов или
капельки бульона) равномерно распределяли по арене,
так что проводить
мобилизацию не
имело смысла, пищу должны
были собирать одиночные фуражиры;
в качестве кормушки с сиропом использовали пипетку,
из которой одновременно
могли
пить
не более пяти муравьев,
что делало бесполезной
мобилизацию и на эту кормушку.
Через один-два месяца, когда муравьи
привыкли к предложенным им режимам,
провели экзамен,
проследив за реакцией
муравьев на кормушку,
выставленную
на арену при свете, т. е. как в контрольной
группе. Вот тут и выяснилось, что
хотя муравьи во всех
группах были
генетически
близкородственными,
но вели они себя совершенно по-разному.
В контрольной группе они проводили
типичную мобилизацию,
обычно не пользуясь
при этом
пахучим
следом,
оставляемым разведчиком
по пути в гнездо. Муравьи второй
группы привыкли
проводить мобилизацию в темноте и
поэтому на экзамене
старательно
шли по проложенному
разведчиком следу, повторяя все его
изгибы, чего в природе мирмики не
делают; нередко муравьи даже добирались
до
кормушки
по следу без разведчика. В третьей
группе
поставленные в жесткие рамки по времени
муравьи научились проводить мобилизацию
значительно
оперативнее, чем
в естественных условиях.
Они бурно реагировали
на первый же приход разведчика, и
уже через несколько минут число муравьев
на кормушке
достигало максимума;
вторичные
мобилизации муравьи обычно
не
практиковали, ведь они привыкли,
что
кормушка всегда исчезает через час.
Ну и, наконец,
муравьи в последних
двух группах
оказались
абсолютно неспособными
провести мобилизацию, поскольку условия
их жизни не требовали такого
умения. Они не воспринимали след
разведчика как сигнал и плохо шли
за лидером,
в результате образующиеся цепочки
муравьев чаще всего распадались,
не достигнув
кормушки.
Результаты этих экспериментов говорят
о том, что мирмики и, вероятно, многие
другие
муравьи
не имеют
жестких
врожденных систем мобилизации,
но способны
создавать их применительно к конкретным
условиям обитания. Несомненно, поведение
при прокладке
следа, движении
по следу, тактильные
и прочие активирующие сигналы основаны
на врожденных механизмах. Но то,
как эти элементы будут
сочетаться друг с другом в системе
мобилизации, зависит от конкретной
ситуации, опре-
360
деляется обучением и приобретением
опыта муравьями-фуражирами. Например,
след
разведчика может играть очень
важную или незначительную сигнальную
роль. Муравьи могут бурно реагировать
на активирующие сигналы разведчика
или почти
не замечать их, могут старательно
бежать цепочкой
за лидером или предпочитать индивидуальную
фуражировку. Мы пока не
знаем, как
происходит такое
обучение, каким образом формируются
различные системы мобилизации.
Для того чтобы
это установить, нужны кропотливые
исследования. Но теперь совершенно
ясно, что
многие коммуникативные системы
у общественных насекомых достаточно
подвижны, лабильны,
могут перестраиваться,
приспосабливаться к окружающим
условиям.
Несомненно, что общественные насекомые
используют способности к обучению,
приобретению опыта
гораздо шире, чем принято
думать. Это позволяет
им эффективно приноравливаться
к различным изменениям среды
обитания, расселяться
и процветать
даже в очень суровых условиях.
Такая
пластичность поведения — одно
из преимуществ социального образа
жизни по сравнению с одиночным.
Иногда на основе приобретения
опыта у общественных насекомых
может
возникать некоторое подобие
«культурных традиций»
— передачи сведений
из поколения
в поколение. Например, у рыжих
лесных муравьев сеть дорог и
тропинок на
территории, размещение
охотничьих
участков
удивительно
стабильны
и сохраняются
практически
неизменными многие годы; тлей
муравьи также разводят всегда
на одних
и тех же деревьях.
Удалось установить,
что эта
стабильность базируется
на
зрительной
памяти муравьев, не утрачивающейся
у
них в течение долгой зимы. Весной
первыми из
муравейника
выходят
самые старые
фуражиры,
которые помнят
все ориентиры
и прочие особенности участков.
Они и восстанавливают дорожную
сеть.
Молодые муравьи, только
приступающие к работам вне
гнезда, постепенно
осваивают
территорию, запоминают ориентиры
и так
приобщаются к «традициям» своей
колонии.
Если отловить с дороги и поверхности
примыкающего к ней сектора купола
всех фуражиров, то
когда через несколько
дней из гнезда
выйдут молодые
рабочие,
они не смогут восстановить прежнюю
дорожную сеть, поскольку не обладают
необходимой
информацией. Но если убрать
только
муравьев
с дороги,
оставив тех, кто находится
на куполе, то муравьи во всех
деталях реконструируют всю систему
дорог.
Оказывается, самые старые
фуражиры остаток
своей жизни
проводят на
куполе в должности
так называемых наблюдателей.
Они несут постоянную вахту и
первыми
сообщают
о приближении врагов.
Кроме того,
они являются
хранителями
«общественных
традиций» — информации об охотничьих
угодьях, удобных для разведения
тлей деревьях и
системе дорог, созданной
предшествующими
поколениями
рабочих.
В экстремальных условиях, когда
исчеза-
361
ли все фуражиры колонны, наблюдатели,
покинув свои посты на куполе,
выходили на территорию
и помогали
«новобранцам»
восстановить
дорожную сеть. Конечно,
подобая «передача традиций»
не предполагает непосредственного
обучения молодых
муравьев старыми, но все
же свидетельствует о социальной
преемственности, основанной
не на биологической наследственности,
а
на приобретении
индивидуального опыта новыми
поколениями насекомых.
Ну, а как же все-таки насчет
разума у общественных насекомых?
Современная
наука
обладает несравненно
большим количеством
экспериментальных данных
по этому вопросу, чем ученые
начала
XX
в. Сразу нужно
сказать,
что
огромное множество фактов
подтверждает значительную
ограниченность
интеллектуальных способностей
общественных насекомых
по сравнению
не только с человеком, но
и с высшими позвоночными.
Очень многое в поведении
муравьев,
пчел, ос, термитов
определяется
врожденными
реакциями
на довольно простые
стимулы
и не предполагает
способностей к сопоставлению
и анализу ситуации в целом.
О каком
разуме
может идти речь,
когда муравей
выносит
из гнезда
на свалку
живого и
здорового собрата
на том лишь основании, что
от него пахнет, как от трупа?
То,
что этот
«живой покойник»
сопротивляется,
вырывается
и вновь
возвращается
в
гнездо, его
непреклонные могильщики игнорируют.
Для них важен только один
признак — запах,
а все
остальное не существует.
Разве способен рассуждать
и сопоставлять факты муравей,
заботливо облизывающий
и снабжающий кормом личинку
жука-атемелеса, которая безнаказанно
пожирает муравьиный расплод?
Не ближе к разумному
и
поведение муравьев, проявляющих
«нежную привязанность» к
кусочку
картона,
смоченному экстрактом личинок,
или пчел, образующих свиту
вокруг ватного тампона, пропитанного
раствором кетодеценовой
кислоты.
Можно продолжить ряд
подобных примеров, когда
насекомые действуют, слепо
подчиняясь
одним стимулам
и абсолютно
не принимая в расчет
все остальные. Все это показывает,
что насекомые
в целом не в состоянии рассуждать,
сравнивать и поступать в
соответствии с
анализом, т.
е. разумно.
Когда в опытах Т. Шнейрлы
муравьям, научившимся хорошо
ориентироваться
в лабиринте, предложили
преодолеть
его в обратном направлении,
они совершенно растерялись
и им пришлось изучать его
заново. То
же самое
получилось
и в
экспериментах известного
русского зоопсихолога В.
Вагнера со
шмелями.
Медоносные пчелы
продемонстрировали ничуть
не лучшие
способности.
В то же время крысы,
например, скоро соображают,
что лабиринт прежний, и на
основе
опыта быстро
находят в нем путь.
Муравьи оказались
еще более
ограниченными.
Когда
они научились проходить через
лабиринт от гнезда к
кормушкеу возвращаясь обратно
по прямому пути, Шнейрла
предложил им
362
вернуться в гнездо по знакомому
уже лабиринту. Муравьи
восприняли эту
ситуацию как целиком
новую и не смогли
воспользоваться своей
памятью о лабиринте
лишь потому, что встретились
с ним в иной обстановке.
А ведь с точки зрения
человека задача эта совсем проста,
здесь
не нужно даже «раскручивать»
память
в обратном
направлении — лабиринт
следовало пройти привычным курсом.
Значит, общественные насекомые
не умеют использовать
накопленный опыт
в новой
ситуации? Да, во
многих случаях для них
это непосильно. Но,
оказывается, не всегда.
В опытах Н. Г. Лопатиной
пчел
обучали
отыскивать корм
в лабораторном
лабиринте, ориентируясь
на свет расположенных
в определенной
последовательности цветных
светофильтров. Затем
обученным пчелам предлагали найти
корм на открытом
воздухе. В этой ситуации
они четко избирали и
входили в тот из четырех боковых
коридорчиков
прибора, в котором были
видны через
стекло кружки
из бумаги
такого же размера и цвета,
расположенные
в той же последовательности,
что и светофильтры при
обучении в лабиринте.
Стало быть, перенос
опыта в новые условия
все-таки иногда
возможен? Жаль, что этому
интереснейшему явлению
пока посвящено так мало
исследований.
Здесь мы подходим к удивительнейшему
факту: наиболее высокоорганизованные
общественные
насекомые —
медоносные пчелы и муравьи,
— несмотря на явное преобладание
в их поведений инстинктов
и интеллектуальную
ограниченность,
все же обладают зачатками
рассудочной деятельности.
Это проявляется
в способности к элементарному
обобщению,
синтезу. В экспериментах
Лопатиной пчел
научили
находить кормушку по
запаху амилового спирта,
а затем
добавили еще
один стимул
— карточку
красного цвета
под кормушкой;
она свидетельствовала,
что кормушка пуста, хотя
и пахнет
амиловым
спиртом. И вот после
15 безуспешных попыток
найти пищу пчелы уловили
значение
красного цвета
и не прилетали на
кормушку,
когда видели под ней
карточку. Но если
карточки не было,
запах амилового
спирта по-прежнему привлекал
их.
Еще поразительнее то,
что красный цвет
приобрел для этих
пчел значение «условного
тормоза»
и в
других ситуациях,
в частности,
когда кормушка пахла
бутиловым спиртом или
была иной формы
и цвета. Получается,
пчелы
сделали
элементарное обобщение:
в присутствии
красного цвета кормушка
всегда пуста.
В других опытах удалось
выработать у пчел представление
о комплексе
стимулов. Например,
они научились
выбирать кормушку
с запахом
октилового спирта
на синем фоне, но отвергали
кормушку с тем же запахом
на белом фоне
и кормушку без запаха
на синем фоне.
Сумели пчелы
запомнить
и различить также последовательности
разных
стимулов. В одном эксперименте
они должны
были подлетать сначала
к синему экрану, затем
к желтому и наконец
к розовому и только тогда
получали
363
корм. Чтобы научиться
делать это, пчелам
потребовалось от 40 до 100
попыток,
но они все-таки решили
задачу!
Когда пчелы научились
различать желтый
и синий треугольники,
опускаясь всегда
на синий,
где
их ждала пища, экспериментаторы
изменили ситуацию
на обратную,
положив подкрепление
на желтый треугольник.
Пчелам
пришлось
переучиваться, и
вскоре они безошибочно
выбирали желтый
цвет. И опять перемена
— подкрепление вновь
на синем треугольнике.
Пчелам
снова
надо переучиваться.
Но вот что интересно:
с каждым разом они
делали
это все быстрее и
быстрее.
Они как бы уловили
закономерность:
если кормушка
пуста, значит, подкрепление
ждет
на соседнем столике.
Такой же способностью
обладают и муравьи.
А. Я. Карась
и М. И. Жуковская
обучали
рыжую мирмику
отыскивать
куколку в симметричном
лабиринте,
она могла быть либо
в правой, либо в
левой его половине.
Когда муравей
привыкал
сразу
приходить, предположим,
в левую половину,
куколку
перемещали
в правую. После
того как он научился
поворачивать безошибочно
направо,
куколку
опять перекладывали
в левую половину,
и т.
д. В
начале опыта муравьям
требовалось большое
время, чтобы
научиться
бежать сразу в противоположную
сторону и
не заглядывать в
симметричную часть
лабиринта. Но
с каждой переменой
муравьи решали
задачу
все быстрее. Некоторые
проявили просто удивительную
сообразительность:
если
куколки не было на
привычном месте,
они сразу же
устремлялись в противоположную
половину лабиринта.
Выходит, пчелы и
муравьи могут создавать
представление
об «альтернативной
ситуации».
В экспериментах московского
энтомолога Г. А.
Мазохина-Поршнякова
медоносные
пчелы продемонстрировали
поразительную
способность к так
называемому инвариантному
опознаванию
фигур и предметов.
Что это означает?
Пчелы
сумели узнавать
геометрические фигуры
вне зависимости от
их краски, формы,
размеров, расположения
в пространстве
и цвета окружающего
фона. Следовательно,
они могут после предварительного
обучения различать
треугольник,
квадрат
или четырехугольник
как таковые, т. е.
как
геометрические фигуры.
Кто бы подумал, что
пчелы столь
способные геометры!
В дальнейших
опытах пчелы показали
еще более удивительные
результаты.
Они,
например, узнавали
фигуры по
признаку двуцветности
вне зависимости от
самих цветов
и формы фигуры
и даже
научились «считать»:
выбирали карточку
с двумя темными
пятнами из таких
же карточек с одним
или тремя пятнами,
хотя
форма, размеры и
взаимное расположение
пятен
во всех повторениях
опыта
были различными.
Немного позже инвариантное
опознавание геометрических
фигур обнаружили
и у кровавого муравья-рабовладельца.
Выходит,
что пчелы и муравьи
могут создавать
элементарные отвлеченные
представления о треугольнике,
круге, квадрате,
коли-
364
честве (в пределах
двух-трех) цветов
или объектов.
На такое способны
только наиболее
высокоорганизованные
позвоночные
животные. Золотые
рыбки, в частности,
не узнают ту
же фигуру, если она изображена
иной краской
или иначе
повернута. Как
поразительно
у общественных насекомых это сочетание
способности к
элементарному
обобщению и быстрому обучению
в одних ситуациях
с крайней
ограниченностью
и невозможностью простейшего
сопоставления
в других, а также с несомненным
преобладанием
врожденных форм поведения!
Высшим интеллектуальным
достижением среди
животных нередко
считают
использование простейших орудий.
Подобное
поведение в некоторых
случаях наблюдали
и
у общественных
насекомых. Рабочие
североамериканских
муравьев рода
афеногастер,
обнаружив
раздавленную
гусеницу, гнилую
ягоду или
другой источник
полужидкой
пищи,
подбирают вокруг
кусочки древесины,
листьев, сухую
грязь и бросают
на добычу.
Когда эти частицы
пропитаются полужидкой
кашицей,
муравьи уносят
их в гнездо,
где тщательно
облизывают и
высасывают
все съедобное.
Употребление
посторонних
пористых предметов
в качестве «сосудов»
позволяет
муравьям
ускорить транспортировку
корма в гнездо.
В зобике муравей
может
принести
в среднем около
0,13 мг пищи,
а в
«сосудах» — 1,38
мг, т. е. в 10
раз больше.
Немаловажно и
то, что такое
поведение дает
возможность «обманывать»
муравьев-конкурентов.
Кампонотусы,
формики и другие
более сильные
муравьи
прогоняют
робких
афеногастеров
с добычи.
И тогда их выручают
примитивные «сосуды».
Подкравшись
к
добыче, муравей
бросает на нее
листик или
кусочек грязи
и тут же ретируется.
А через
минуту-две тихонечко
возвращается
и
уносит свой «сосуд»
с
желанной провизией
в гнездо. Набрать
корм в зобик
в присутствии
враждебных муравьев
никак
невозможно, а
с
помощью «сосудов»
афеногастеры
воруют корм из-под
самого носа конкурентов.
Рабочие завезенного
в США дернового
муравья охотятся
на пчел-галиктов,
охраняющих
входы в норки,
бомбардируя
их
мелкими камешками.
Без этого пчелу,
закрывающую отверстие
своей «бронированной»
головой, не выманить.
Муравьи собирают
вокруг
камешки и
сбрасывают
в норку на голову
сторожа. Такая
наглость крайне
злит
пчелу. Сначала
она пытается
прогнать муравьев,
раскрывая челюсти,
выставляя вперед
брюшко с обнаженным
жалом,
но бомбардировка
продолжается.
Тогда потерявшая
всякое
терпение пчела
выходит из норки,
и
тут на нее набрасывается
целая
орда муравьев,
которые только
этого и ждали.
Теперь участь
пчелы печальна.
Вот как подводит
пчелу невыдержанность,
а муравьям помогает
их
«оружие»! Обитающие
в пустынях
Аризоны
муравьи кономирма
биколор аналогичным
образом
борются с конкурентами.
Они осаждают
находящиеся
неподалеку гнезда
медовых муравьев,
муравьев-жнецов
и непрерывно
бросают
во входы камешки,
вынуждая
365
обитателей прекратить
фуражировку
и забаррикадироваться
в
гнезде.
Конечно,
орудийная
деятельность
муравьев
очень проста,
но все же
представляет
для нас огромный
интерес.
Являются
ли
эти действия
врожденными,
или муравьи
обучаются
им, тогда
каким
путем? Используют
ли орудия
все
муравьи данного
вида или
лишь некоторых
популяций?
На
эти вопросы
у нас пока
нет ответов.
Будем надеяться,
что
их принесут
новые исследования.
Таким образом,
хотя наука
обладает
солидным
багажом
экспериментальных
данных об
интеллектуальных
способностях
общественных
насекомых
и об
их поведении
мы
знаем
гораздо больше,
чем сто лет
назад, надо
признать,
что наши
общие, теоретические
представления
по этой
проблеме
недалеко
ушли
вперед. И
сегодня вполне
современно
звучит точка
зрения О.
Фореля, К.
Эмери и многих
других ученых
прошлого
о
том, что
поведение
общественных
насекомых
в основном
инстинктивно,
но у
них
имеются элементы
разумной
деятельности.
Смогут ли
будущие поколения
ученых
прибавить
к этой формулировке
что-нибудь
существенное?
ФЕНОМЕН КОЛЛЕКТИВА
Пойди к муравью, ленивец, посмотри на действия его и будь мудрым. Нет у него ни начальника, ни приставника, ни повелителя, но он заготовляет летом хлеб свой, собирает во время жатвы пищу свою... Книга притчей Соломоновых Что же такое этот «дух улья», где он находится?.. Он безжалостно располагает здоровьем и счастьем, свободой и жизнью всех этих крылатых созданий и вдобавок с благоразумием, будто управляет самим собою с сознанием величайшего долга. М. Метерлинк. Жизнь пчел |
Поведение отдельного муравья, пчелы или термита нередко может быть очень сложным, особенно, если оно связано с приобретением опыта или элементарной рассудочной деятельностью. Но все-таки наиболее интересны общественные насекомые тем, на что они способны все вместе. В большинстве случаев каждое насекомое выполняет лишь ничтожную часть общего дела, приводящего к результату вследствие суммирования усилий десятков, сотен и даже тысяч его собратьев. Благодаря коллективной деятельности насекомые строят удивительные по архитектуре гнезда, регулируют в них микроклимат, запасают пищу, зачастую специально перерабатывая и даже консервируя ее, оборо-
366
няются
от
врагов, осваивают
и
охраняют
от
конкурентов обширные
кормовые
угодья,
расселяются
и
порой мигрируют
на
значительные расстояния.
Как
организовано коллективное
поведение насекомых?
Как
на основе
интеграции
действий
множества
отдельных
особей
достигается
целесообразный
результат?
Кто
управляет ими?
Из
глубокой
древности
идет
сохранившееся
до
наших
дней
представление
о
бесконечном трудолюбии
пчел
и
муравьев, с готовностью
жертвующих
личным
благополучием
и
отдающих все
силы
ради
общественного
блага.
П.
Латрейль
в
книге «Естественная
история
муравьев»
ярко
выразил
этот
традиционный
взгляд:
«Можно
ли
найти
другое
такое
общество,
члены
которого
обладали
бы
большей
любовью
к
окружающим? Были
бы
более
заинтересованными?
Проявляли
ли
бы
в
работе
столь
упорное
и
неослабное рвение?
Какой
уникальный
феномен!»1
Не
менее
распространено
и
мнение
об
исключительно
рациональной
организации
общества
насекомых,
действующих
всегда
согласованно,
целесообразно
и
в соответствии
с
коллективными нуждами.
Однако
наука
постепенно
развенчала
эти
красивые
мифы.
Оказалось,
что
дело
обстоит
не
так.
Чтобы
лучше
понять
это,
приглядимся
внимательней
к
коллективной деятельности
общественных
насекомых
на
нескольких
примерах.
Вот
муравьи
переселяются
из
ставшего
непригодным
старого
гнезда.
Разведчики
уже
нашли
подходящее
место,
проложили
туда
пахучую
тропинку
и
провели
мобилизацию
в
гнезде.
Живой
ручеек
из
муравьев
постепенно
перетекает
к
месту
новоселья.
Многие
переселенцы
несут
яйца,
личинок,
куколок,
молодых
рабочих.
Доставив
груз
в
новое
гнездо,
они
возвращаются
за
следующей
ношей.
Но
что
это?
Некоторые
муравьи
тащат
тот
же
груз
в
обратном
направлении,
в
старый
муравейник,
а
затем
опять
и
опять
возвращаются
за
личинками
и
куколками,
только
что
размещенными
в
новом
гнезде
другими
носильщиками,
чтобы
водворить
их
на
прежнее
место.
Их
действия
явно
не
координируются
с
поведением
остальных
муравьев
и
мешают
скорейшему
достижению
общей
цели.
Есть
в
потоке
муравьев
и
такие,
что
бегают
безо
всякой
ноши
туда
и
обратно.
Даже
если
принять
во
внимание
необходимость
дополнительного
усиления
пахучего
следа,
все
равно
количество
порожних
рейсов
столь
велико,
что
никак
не
согласуется
с
представлением
об
исключительной
целесообразности
происходящего.
В
конце
концов
муравьев,
конечно,
можно
будет
поздравить
с
новосельем:
весь
расплод
и
все
взрослые
окажутся
в
новом
гнезде.
Причина
этого
проста:
муравьев,
таскающих
расплод
в
обратном
направлении,
немного.
Но
так
ли
уж
рационально
было
организовано
переселение?
1 Latreillе P. A. Histoire naturelle des fourmis. Paris, 1802. P. 237.
367
Попробуем
теперь в
процессе переселения
муравьев из
одного лабораторного
формикария в
другой открыть
доступ свету
в новое
помещение, уменьшить
в нем
влажность или
как-нибудь иначе
ухудшить «жилищно-бытовые
условия». Количество
муравьев, таскающих
расплод в
обратном направлении,
сразу возрастет,
и на
некоторое время
может установиться
равновесие между
числом приносимых
и уносимых
куколок и
личинок. Если
же обстановка
в новом
помещении окажется
еще хуже
и не
будет устраивать
многих муравьев,
переселение повернет
вспять и
вскоре все
вернутся в
прежнее гнездо.
Значит, все
решает большинство
и процесс
организован по
принципу плебисцита
— всеобщего
и равного
голосования. Естественно,
что из
«голосования» исключены
«несовершеннолетние» —
расплод и
молодые муравьи,
которых транспортируют
носильщики; царица
тоже подчиняется
большинству, как
это и
должно быть
при конституционной
монархии!
Аналогичным
образом происходит
выбор места
для поселения
роем медоносных
пчел. Вспомним
«диспут пчел-квартирмейстеров»,
во
время которого
пчелы «принимают
решение большинством
голосов» и
перелетают туда,
куда направляет
их основная
масса танцующих
на поверхности
роя разведчиц.
В одном
опыте М.
Линдауэра семью
пчел перевезли
на маленький
и пустынный
островок в
Балтийском море.
Когда началось
роение, экспериментаторы
поставили на
разных концах
острова два
одинаковых пустых
улья. Это
привело пчел
в совершенное
замешательство. Поскольку
оба помещения
были равноценными,
примерно половина
квартирмейстеров танцами
указывала пчелам
на один
улей, а
другая половина
не менее
усердно призывала
их поселиться
во втором.
Так продолжалось
три дня
— и
пчелы все
не могли
выбрать себе
жилище. В
конце концов
рой взлетел,
но разделился
на две
части и
каждая направилась
к своему
улью, возглавляемая
когортой неуступчивых
квартирмейстеров. В
естественных условиях
никогда не
бывает абсолютно
одинаковых помещений
и поэтому
система «демократического
выбора» пчел
не испытывает
затруднений.
Вернемся
опять к
муравьям и
попытаемся выяснить,
насколько рационально
организовано у
них, например,
кормление личинок.
Для этого
возьмем 10
личинок и
дадим им
одного муравья-няньку.
Взвесив личинок
до опыта
и, допустим,
через два
часа, мы
узнаем, сколько
корма получили
они от
няньки. Другой
группе личинок
выделим двух
нянек, третьей
— трех
и т.
д. Когда
все личинки
будут взвешены,
выявится очень
любопытная картина.
Пока нянек
немного (2—4),
вклад каждой
в кормление
личинок примерно
одинаков. Но
когда их
больше (5—7),
средний привес
личинок в
пересчете на
одного муравья
становится меньше
и продолжает
снижаться
при
дальнейшем росте
числа нянек.
Значит, каждая
нянька в
среднем дает
личинкам меньше
пищи. Почему
же так
происходит?
368
Дело
в том,
что муравьи
кормят личинок
по методу
«проб и
ошибок». Рабочий
заранее не
знает, какая
из них
готова принять
пищу, и определяет это
во время
тактильного контакта
с личинкой,
постукивая антеннами
по ее
ротовым органам.
Когда нянек
немного, каждая
может встретить
голодную личинку
с большей
вероятностью, чем
только что
накормленную, и
поэтому почти
любая ее
попытка кормления
окончится успешно.
Но если
нянек больше,
они значительно
чаще будут
предлагать корм
сытым личинкам,
не желающим
некоторое время
питаться. Коэффициент
полезного действия
каждого муравья
будет в
этих условиях
меньше. В
результате все
личинки получат
необходимое им
количество пищи,
но сколько
усилий муравьев-нянек
окажется растраченным
впустую! Можно
ли после
этого считать
организацию кормления
личинок у
муравьев исключительно
рациональной?
Эффективность
рабочих уменьшается
не только
потому, что
они напрасно
дублируют друг
друга, есть
и иные
причины. Личинки
в гнезде
нередко сложены
кучками. Это затрудняет нянькам
доступ к
тем, кто
находится внизу,
и уменьшает
продуктивность их
деятельности. Недаром
муравьи стараются
размещать личинок
в камерах
гнезда как
можно более
равномерно. Но
далеко не
всегда это
у них
получается. Вот
почему система
выращивания расплода
у пчел
и ос,
при которой
каждая личинка
занимает отдельную
ячейку, значительно
рациональнее, чем
у муравьев.
Вторая
причина неэффективности
рабочих связана
с их
численностью и
распределением. Если
муравьев в
данном месте
слишком много,
то они
мешают друг другу, затрудняют
доступ к
личинкам и
коэффициент их
полезного действия
снижается. Так
происходит не
только при
кормлении личинок,
но и
в иных
случаях. Например,
когда муравьи
роют ход,
количество выкапываемой
в пересчете
на одного
рабочего почвы
максимально при
3—5 работающих.
При большем
количестве землекопов
их индивидуальная
выработка намного
уменьшается, поскольку
они путаются
под ногами
друг у
друга. Получается,
что реальный
вклад рабочих
в общественное
благосостояние зависит
от очень
многих факторов
и далеко
не всегда
так высок,
как можно
было бы
предполагать.
Рассмотрим
еще один
пример —
строительство воскового
сота медоносными
пчелами. Если
внимательно приглядеться
к происходящему,
то прежде
всего бросается
в глаза хаотичность, беспорядочность
действий
отдельных пчел.
Вот строительница
прилепила к
краю сота
кусочек воска,
придала ему
определенную форму
и отошла
в сторону.
Другая пчела,
появившаяся на
этом месте,
тут же
оторвала кусочек
и пристроила
его неподалеку.
То же
самое совершается
и вокруг:
одни строят,
другие
ломают
и переделывают
на свой
лад. Пчелы
действуют в
полном смысле
слова наперекор
друг другу.
Но поскольку
строят
369
они
все же
больше, чем
разрушают, сооружение
постепенно увеличивается
и вот
— о
чудо! —
перед нами
великолепно изготовленный, геометрически правильный
сот. Очень
похожую картину
строительных работ
можно увидеть
в гнездах
ос, шмелей,
термитов —
любых общественных
насекомых. Как
же возникает
порядок из
хаоса?
Прежде
всего из
уже сказанного
ясно, что
в колонии
общественных насекомых
нет какого-либо
управляющего центра,
механизма, регламентирующего
деятельность каждого ее
члена. Если
регуляция и
осуществляется, как,
например, в
случае репродуктивного
контроля царицы,
то она
носит исключительно
общий характер
и не
затрагивает конкретной
деятельности отдельных
насекомых. Коллективное
поведение формируется
путем суммирования
действий множества
индивидуумов, т.
е. является
стохастическим
процессом.
При этом
отдельные насекомые
могут действовать
неправильно, нерационально,
вхолостую и
даже во
вред коллективу.
Но поскольку
большинство прилагает
усилия в
определенном направлении,
при усреднении
возникает целесообразный
результат. Следовательно,
наблюдающийся порядок
является исключительно
статистическим. В
этом и
заключается феномен
коллективного поведения
общественных насекомых.
Хорошей
его иллюстрацией
служит прокладка
пахучей тропы
термитами или
продвижение вперед
фуражировочных колонн
муравьев —
жнецов и
кочевников. По
наблюдениям Т.
Шнейрлы, перемещение фуражировочного скопления
эцитонов определяется
двумя процессами.
Во-первых, муравьи
стараются покинуть
участки со
слишком высокой
плотностью особей.
Поскольку сзади
по тропам
подходят все
новые контингенты
рабочих с
бивуака, фуражиры
в скоплении
вынуждены постепенно
перемещаться вперед.
Во-вторых, как
только муравьи,
продвигаясь, освобождают
хоть какое-то
пространство, его
тут же
заполняют рабочие
из соседних
участков с
повышенной плотностью;
этот механизм
обеспечивает непрерывность,
связанность фуражировочного
скопления. Его
фронт продвигается
вперед благодаря
челночным движениям
множества прокладывающих
следы муравьев.
Хотя траектории
движения особей
хаотичны, фуражировочное
скопление перемещается
как единое
целое в
определенном направлении.
Вероятно,
одним из
первых понял
суть коллективного
поведения общественных
насекомых создатель
кибернетики Н.
Винер. Согласно
его идее,
колония общественных
насекомых состоит из элементов
(особей) с
простым стереотипным
поведением, а
сложность ее
коллективной деятельности
определяется связями
и взаимодействиями
между элементами.
Но от
подобного понимания
еще очень
далеко до
настоящего объяснения.
Попытаемся хотя
бы немного
прояснить эту
проблему.
Если
внимательно понаблюдать
за поведением
отдельных пчел
или муравьев
внутри гнезда,
хронометрируя их
деятель-
370
ность
за длительный
период, легко
убедиться в
том, что
более половины,
а нередко
и свыше
двух третей
всего времени
насекомые заняты
«ничегонеделанием» — бродят без
видимой цели
по гнезду
или пребывают
в неподвижности,
как бы
«отдыхают». Эта
картина не
очень-то согласуется
с тезисом
о всепоглощающем
трудолюбии общественных насекомых! Но
свидетельствует ли
она об
их лености?
До
сих пор
неизвестно, необходим
ли насекомым
отдых как
средство восстановления
сил и
есть ли
у них
нечто подобное
сну высших позвоночных. Поэтому
трудно сказать,
отдыхают ли
пчелы или
муравьи, когда
неподвижно сидят
в гнезде.
Возможно, отчасти
это действительно
так. Но
все-таки основное
значение пассивного
времяпрепровождения, видимо,
состоит
в том,
что оно
позволяет насекомому,
не находящему
применения своим
силам, не
тратить впустую
энергию пищи,
являющейся достоянием
всей колонии.
Поэтому, когда
нет дел,
в гнезде
«отдыхают» и
фуражиры, и
строители, и
рабочие других
функциональных групп.
Мы уже
говорили о
наличии в
гнездах общественных
насекомых так
называемых резервистов
— длительный
период ничем
не занятых
рабочих. Резервисты
вместе с
временно неактивными
насекомыми составляют
запасную «армию
труда», которая
при потребности
может быть
мобилизована на
те или
иные работы.
Но если
«отдыхающих» фуражиров
направляют за
добычей разведчики, то внутригнездовые
рабочие сами
озабочены поисками
себе дела.
Для
этого они
и бродят
туда-сюда по
гнезду. Эти
«прогулки», которые
М. Линдауэр
назвал патрулированием,
несмотря на
внешнюю праздность,
имеют определенную цель —
поиски работы.
Проследим за
поведением отдельной
пчелы в
улье. В
«Кастах и
разделении труда»
мы уже
говорили о
том, что
пчелы в
улье выполняют
различные функции
без какой-либо
четкой последовательности.
Вот пчела
идет по
соту, обнаружила
покинутую молодой
пчелой ячейку,
вычистила ее,
отправилась дальше.
Патрулирование продолжается
некоторое время.
Иногда его
прерывают короткие
периоды «отдыха».
А когда
снова найдено
дело, пчела
ремонтирует поврежденный
сот, кормит
личинку, запечатывает
ячейку с
куколкой или
запасом корма.
И опять
пчела патрулирует
в поисках
работы. Никто
не сообщает
пчеле, где
нужна ее
деятельность, никто
не направляет
ее в
тот или
иной конец
улья. Она
сама находит
применение своим
силам. Значит,
патрулирование ничего
общего с
бездельем не
имеет, а представляет собой
форму организации
работ у
общественных
насекомых.
Теперь
мы можем
сделать очень
важный вывод.
Своевременное и
быстрое выполнение
всех дел
в гнезде
общественных насекомых
обеспечивается отнюдь
не тем, что они
детально согласуют
между собой
свои действия
или тщательно
планируют работы.
Напротив, насекомые
трудятся практически
независимо
371
друг
от друга.
Но в
гнезде, как
правило, существует
большой избыток
непрерывно отыскивающих
занятие насекомых
и благодаря
этому все
необходимые дела бывают выполнены
незамедлительно. Можно
сказать, что
эффективное ведение
хозяйства здесь,
как и
в капиталистических
странах, обеспечивает
временная или
постоянная «безработица»
значительной части
насекомых! Можно
ли назвать
подобную систему
наиболее рациональной?
Видимо, нет.
Но как
бы то
ни было,
факт остается
фактом: одним
из основных
принципов организации
коллективного поведения
у общественных
насекомых является
избыточность
трудовых
ресурсов и
самостоятельный
поиск рабочими задач,
которые требуется
выполнить. Альтернативным
путем организации
работ служит
мобилизация, но
ее используют
далеко не
все виды
и преимущественно
во внегнездовой
деятельности. Таким
образом, на
«бирже труда»
общественных
насекомых
существует
два
главных
метода
повышения
занятости:
индивидуальное
и
самостоятельное
«трудоустройство»
в
той
же
местности
и
«коллективные
контракты» на
работу
в
отдаленных
районах.
Мы
уже
неоднократно
подчеркивали,
что
во
многих
случаях
общественные
насекомые
действуют
практически
независимо
друг
от
друга.
Эту
фундаментальную
особенность их
поведения
впервые
понял
П.
Гюбер.
В
своей
книге
«Исследования
нравов
местных
муравьев»
(1810),
рассказывая
о
строительстве
гнезда
бурым
лесным
муравьем,
он
писал:
«На
основании
этих
наблюдений и
тысячи подобных
им я
убедился, что
каждый муравей
действует независимо
от своих
товарищей. Первый
муравей, который
изобретает легкий
план
выполнения
работы, сейчас
же начинает
его осуществлять;
другим остается
только продолжать
начатое, и
они решают,
что им
следует предпринять,
на основании
осмотра первых
результатов
работы...»2 Однако
этот
глубокий вывод
Гюбера, не
понятый его
современниками, надолго
забыли. Явление
было переоткрыто
почти через
150 лет
известным французским
энтомологом П.
Грассе. Он
пытался отыскать
пути к
решению труднейшего
вопроса: как
удается муравьям,
термитам, пчелам
строить огромные
и очень
сложные гнезда,
если каждое
насекомое может
внести лишь
небольшой вклад
в работы
и не
в силах
даже «окинуть
взглядом» все
сооружение? Короче
говоря, у
кого находятся чертежи гнезда?
Грассе
изучал строительное
поведение африканских
термитов родов
кубитермес и
макротермес в
крайне простых
условиях. Определенное
количество термитов
заключали в
небольшой контейнер с
почвой
и наблюдали,
что они
будут делать.
Сначала термиты
в возбуждении
бегали в
разных направлениях,
обследуя помещение,
затем начали
строить. Но
на первом этапе
в
их деятельности
царил полный
хаос: они
смачивали экскрементами
2 Huber P. Recherches sur les moeurs des fourmis indigenes. Paris; Geneve, 1810. P. 295.
372
комочки почвы, несли и пристраивали их кто куда сможет по всей площади контейнера, нередко один и тот же комочек несколько раз перетаскивали с места на место разные термиты. Но вот случайно термиты прилепили два-три комочка друг на друга. Это маленькое возвышение сразу же оказалось очень привлекательным для насекомых. Наталкиваясь на него, они пристраивали здесь свои комочки и быстро отправлялись за новыми порциями. Так постепенно вырос столбик. В случае, когда комочки первоначально лежали цепочкой, термиты сооружали

86. Строительство арки термитами макротермес наталензис (по Э. Вильсону, 1971 г.).
Рабочие
сооружают колонны
из комочков
почвы и
экскрементов. Одни
прилепляют
строительный
материал сбоку,
другие прикрепляют
комочки на
вершине. Затем
строители
начинают изгибать
колонну по
направлению к
соседней. В
результате получается
арка,
показанная на
заднем плане.
на этом месте вал, форма которого определялась конфигурацией цепочки. Если поблизости не было подобных конструкций, термиты прекращали строительство и переходили на иные участки. Но когда по соседству шли такие же работы, строители продолжали приносить новые комочки почвы, надстраивая свой столбик или вал. Едва их высота достигала определенной величины, как поведение термитов вновь менялось. Теперь они приклеивали строительный материал не в центре столба, а ближе к краю, постепенно загибая его вершину по направлению к бли-
373
жайшей
колонне. Их
соседи поступали
точно так
же. Поэтому
вскоре обе
изогнутые колонны
соединились, образовав
арку. Два
соединенных соседних
вала становились коридором. Мало-помалу
термиты объединяли
и ближайшие
арки так,
что возникала
система тоннелей
и навесов,
в которых
скрывались от
внешнего мира
маленькие строители.
До
сих пор
неизвестно, как
удается слепым
термитам в
полной темноте
совмещать концы
обеих половинок
арки наверху.
Возможно, они
ориентируются по
запаху, хотя не исключено,
что они
руководствуются электростатическими
полями,
существующими вокруг
тела любого
насекомого. В
данный момент
для нас
это не
имеет значения.
Важно уловить
основной принцип
организации строительства.
Он заключается
в том,
что термиты,
сообразуя свое
поведение с
уже построенным,
продолжают работу предшественников.
Каждый
термит ведет
себя так,
будто
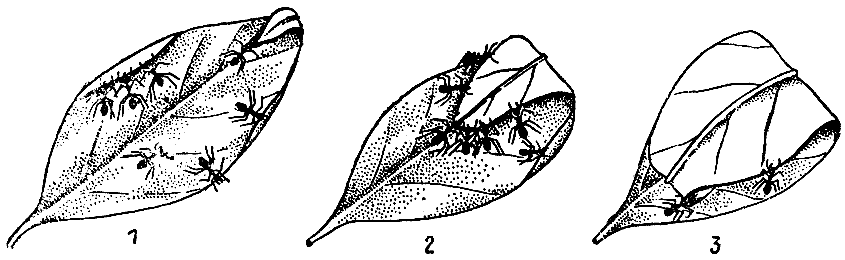
87. Принцип стигмэргии в строительной деятельности муравьев-портных (по Дж. Садду, 1963 г.).
1
— начало
сворачивания
листа;
2 —
все рабочие
присоединились
к
муравью,
достигшему
наибольшего
успеха
на вершине
листа; 3
— совместное
завершение
работы.
он
трудится
один
в соответствии
со своими
врожденными
навыками.
Но поскольку
все термиты
обладают
одинаковыми
или, по
крайней мере,
очень сходными
наследственными
программами поведения,
они
создают общими
усилиями
определенную
структуру.
Можно
в какой-то
момент убрать
из контейнера
всех термитов,
заменив их
новыми. И это
ничего
не изменит:
новички
окажутся
достойными
преемниками
и будут
в основном
продолжать
начатое
их предшественниками.
Грассе
назвал
такой
способ
организации
строительства
принципом
стигмэргии,
в
переводе
с
греческого
это
означает
«побуждение
к работе».
Суть его
заключается
в том, что
результаты работы
одних насекомых
могут служить
стимулом для
последующей деятельности
других. Как
показывают внимательные
наблюдения за
строительным поведением
муравьев, ос,
пчел, и
у них
оно основано
на этом
же принципе.
Например, муравьи-ткачи
начинают сближать
и сворачивать
листья сразу
во многих местах. Но
как только
в какой-либо
точке достигнут
существенный успех,
окружающие муравьи
бросают участки,
где они
374
трудились,
и присоединяются
к более
результативным строителям,
что позволяет
быстро завершить
работу.
Стигмэргическая
организация деятельности
принципиально отличает
общественные виды
от одиночных.
Еще Ж.
Фабр установил,
что поведение
насекомых представляет
собой цепь действий, последовательность
которых
жестко фиксирована.
Пчела-каменщица сначала
строит ячейку,
затем носит
в нее
пыльцу и
нектар, а
потом откладывает
яйцо. Если
ей предложить
уже наполненную
провизией ячейку,
то она
не отложит
яйцо сразу,
а будет
еще некоторое
время приносить
в нее
излишний в
данном случае
корм. Возникновение
социального образа
жизни требует
формирования поведения
нового типа
с нефиксированными
последовательностями действий.
На
это обстоятельство
впервые обратил
внимание ученых
немецкий энтомолог
Г. Легеви
в 1925
г. Например,
уже у
самых примитивных
в социальном
отношении пчел-галиктов
каждая особь
может включиться в общую
работу на
любом ее
этапе, продолжая
то, что
начато другими.
Конечно,
один принцип
стигмэргии не
в силах
объяснить весь
процесс строительства
обширного гнезда.
Для того
чтобы это
сделать, нужно
по меньшей
мере выяснить, от чего
зависят правила
работы, почему
они неодинаковы
в разных
частях «стройплощадки».
Ведь не
всегда же
термиты, например,
возводят столбики
и арки,
в других
ситуациях они
ведут себя
иначе. Неясно
также, отчего
начинается строительство
именно в
том, а
не в
ином месте,
и почему
оно заканчивается
через некоторое
время. Мы
не знаем,
какие особенности
общественных насекомых
определяют значительные
различия в
строении гнезд
нередко даже
очень близких
видов. Мы
не можем
сказать, почему
в зависимости
от внешних
условий муравьи,
термиты и
другие насекомые
могут сооружать
разные гнезда.
В последние
годы появились
попытки компьютерного
моделирования процессов
строительства у
общественных насекомых.
Но это
еще только
начало, и
ответить на
поставленные выше
и многие
другие вопросы
предстоит в
будущем. Хотя
дать ответ
на самый
первый вопрос
мы уже
в состоянии:
чертежи будущего
гнезда находятся
у каждого
строителя. У насекомых
нет
инженеров и
прорабов, любое
из них
строит в
соответствии с
врожденными «чертежами»,
ориентируясь на
то, что
уже сооружено.
Стигмэргия
является одним
из важнейших
принципов организации
коллективного поведения
общественных насекомых
не только
при строительстве,
но и
во многих
иных формах их
деятельности.
Мы уже
неоднократно повторяли,
что общественные
насекомые нередко
действуют совершенно
независимо друг
от друга,
сообразуясь с
ситуацией. Именно
такая деятельность и
соответствует
принципу стигмэргии.
Насекомые кормят
личинок, обрабатывают
принесенную провизию,
выбрасывают мусор
из гнезда
и занимаются
многими иными
делами, сообразуясь
375
в
основном с
объектом своей
деятельности и
не нуждаясь
в получении
какой-либо информации
непосредственно от
собратьев.
Вот
еще один
пример. Высыпем
на открытую
арену муравьев
вместе с
расплодом. Вначале
там будет
царить полная
неразбериха. Муравьи
станут таскать
личинок туда-сюда, складывая их
в разных
местах без
всякого порядка.
Но как
только где-нибудь
случайно появятся
небольшие кучки
расплода, они
сразу окажутся
организующими элементами,
обладающими стигмэргическим
действием. Муравьи
начнут носить
расплод в
эти кучки,
причем нередко
одни будут
притаскивать, а
другие забирать
личинок и
куколок и
транспортировать в
«свою» кучку.
Постепенно в
результате всех
этих несогласованных
действий муравьи
соберут весь
расплод в
несколько кучек.
Но те,
что крупнее,
окажутся привлекательнее,
и сюда
перенесет личинок
и куколок
большее число
муравьев. Поэтому
в конце
концов расплод,
как правило,
сосредоточится в одном месте,
и муравьи
успокоятся, прикрывая
его своими
телами.
Конечно,
принцип стигмэргии
далеко не
всегда лежит
в основе
коллективного поведения
общественных насекомых.
Нередко в
его организации
ведущая роль
принадлежит механизмам
коммуникации, многие
из которых
мы уже
рассматривали. Привлечение,
сигнализация об
опасности, мобилизация
позволяют общественным
насекомым действовать
достаточно согласованно
во имя
достижения какой-либо
цели. Огромное
значение в
управлении коллективным
поведением имеет
так называемая
массовая
коммуникация,
представление о
ней сформулировал
в 1962
г. известный
американский биолог
и мирмеколог
Э. Вильсон.
Массовая коммуникация
по сути
своей коллективна
и заключается
в распространении
внутри группы такой
информации,
которую не
могут сообщить
друг другу
отдельные насекомые.
Чтобы понять
сущность этого
явления, рассмотрим
несколько примеров.
При
мобилизации по
следу количество
прибегающих к
кормушке муравьев
обычно тем
больше, чем
обильнее и
доступнее пища,
когда же
она иссякает,
уменьшается и поток курсирующих
по тропе
фуражиров. Каким
образом происходит
подобная регуляция
фуражировки? Это
впервые было
исследовано Вильсоном
на примере
огненного муравья.
Рабочий, возвращающийся
от кормушки
в гнездо,
может оставлять
за собой
пахучий след
или не
делать этого.
Если пищи
много и
фуражир быстро
наполнил зобик,
то он
проложит след
на всем
пути к
гнезду. Но
если он
долго не
мог протолкнуться
к окруженной
плотным кольцом
муравьев кормушке
или с
трудом заполнил
зобик, слизывая
крохи сахарного
сиропа, то
следа не
оставит и,
скорее всего,
больше сюда
не придет.
Поэтому в
начальный период
мобилизации, когда
пищу брать
легко, пахучий
след усиливается,
привлекая новый
контингент
фуражиров.
Вскоре количество
фуражиров становится
достаточным
для
эксплуатации
данного
источника корма.
Вновь появившиеся
у кормушки
рабочие попадают
в не-
376
выгодное
положение, поскольку
к пище
не могут
проникнуть, и
возвращаются в
гнездо, не
оставляя следа.
Наступает равновесие,
при котором
сохраняется интенсивность
следа, обеспечивающая
эксплуатацию кормушки
примерно постоянным
числом фуражиров.
По мере
того как
количество пищи
убывает, все
меньше возвращающихся
муравьев оставляют
за собой
след, а
многие прокладывают
след лишь
на. некоторых
участках пути
или сокращают
длину и
частоту меток
феромоном. Движение
по тропе
становится менее оживленным и
совсем прекращается
по исчерпании
запаса корма.
В
данном случае
каждый отдельный
муравей ничего
не сообщает
собратьям. Определенная
интенсивность следа
создается в
результате суммирования
зачастую совершенно
различных, реакций множества муравьев,
т. е.
чисто статистически.
Получается, что
информация о
количестве и
доступности корма
существует и
на ее
основе происходит
довольно точная
регуляция фуражировки,
хотя отдельные
муравьи этой
информацией не
обладают и
непосредственно не
передают ее
собратьям. У
других видов
муравьев в
массовой коммуникации при мобилизации
по следу
участвуют еще
тактильные активирующие
сигналы: чем
больше количество
и доступность
корма, тем
интенсивнее проводят
активацию новичков
возвращающиеся в
гнездо фуражиры.
Второй
пример касается
деятельности пчел,
доставляющих в
улей воду
для разбавления
меда и
охлаждения сотов.
Если потребности
в воде
возрастают, например
при перегреве
улья, водоносы
начинают трудиться усерднее и
даже мобилизуют
танцами помощников.
Но когда
воды нужно
меньше, многие
особи прекращают
полеты, невольно
пребывая в
безделье на
сотах, пока
их услуги
вновь не
понадобятся семье.
Исследования М.
Линдаузра показали,
что поведение
водоносов определяется
тем, насколько
легко им
удается сбыть
принесенную в
улей воду.
Если потребность
велика, к
возвратившемуся водоносу
тут же
подскакивает две-три
пчелы, с
жадностью забирающие
у него
воду. Но
когда сильной
«жажды» у
ульевых пчел
нет, водоносу
приходится долго
бродить по
сотам в
попытках пристроить
принесенный груз.
Оказалось, что
если водоносы
затрачивают на
«сбыт воды»
более минуты,
то интенсивность
их полетов
начинает снижаться.
Но если
воду забирают
максимум через
сорок секунд,
водоносы прилежно
трудятся сами
и почти
каждый раз
танцуют, мобилизуя
других пчел
себе на
подмогу. Такая
зависимость приводит
к тому,
что частота
вылетов водоносов
возрастает при
уменьшении среднего промежутка времени
от их
возвращения до
передачи воды
ульевым пчелам.
Можно
подумать, что
при этом
каждый водонос
точно осведомлен
о потребностях
семьи в
воде. Но
нет, на
деле все
совсем не
так и многое
зависит
от случая.
Один водонос
отдает свою
ношу сразу
же по
прилете в
улей, а
другой, возвратившийся
377
вместе
с ним,
долго не
может избавиться
от нее.
Значит, первая
пчела должна
«подумать», что
семью обуяла
непомерная жажда,
а вторая
«решить», что вода не
нужна. На
самом деле
не права
ни та,
ни эта.
Информация об
истинной потребности
в воде,
заключенная в
среднем промежутке
времени, необходимом
водоносам для
сбыта принесенной
воды, не
может быть
достоянием отдельного
насекомого. Эта
информация распространяется
только при
взаимодействии групп
насекомых, передается
от ульевых
пчел водоносам,
тонко управляя
вылетами последних.
Лишь в
предельных ситуациях
— при
очень сильном
перегреве или
полном отсутствии
потребности в воде,
когда
все водоносы
либо работают
в максимальном
темпе, либо
отдыхают, можно
считать, что
они обладают
истинной информацией
о положении
дел.
По такой же
схеме у
общественных насекомых
происходит регуляция
фуражировки, особенно,
если речь
идет о
жидкой пище,
передаваемой фуражирами гнездовым
рабочим
путем трофаллаксиса.
Чем быстрее
в среднем
фуражиры отдают
принесенную ими
пищу, тем
усерднее они
добывают ее,
тем активнее
проводят мобилизацию
новичков. Массовая коммуникация
управляет
коллективным поведением
насекомых часто,
но лишь
в немногих
случаях эти
процессы хорошо
изучены.
Все сказанное выше
позволяет нам
подчеркнуть еще
одну важнейшую
особенность коллективного
поведения общественных
насекомых. Как
правило, его
организация такова, что отдельное
насекомое не
в состоянии
оценивать ход
различных процессов
в колонии
и не
имеет о
них представления.
Муравей, пчела
или термит
не могут
знать, как
много в
их гнезде
личинок, каковы
запасы и
потребности в
пище, сколько
необходимо воды
или строительных
материалов и
т. д.
Они просто
реагируют на
те конкретные
ситуации, в
которые попадают,
и на
те стимулы,
с которыми
встречаются, в
соответствии с
врожденными программами
поведения, приобретенным
в течение
жизни опытом,
физиологическим состоянием
организма и
информацией, полученной
от собратьев
по гнезду.
Организация всех
этих процессов
такова, что
при суммировании,
усреднении нередко
противоречивых, бессмысленных
и
нерациональных действий
множества насекомых
возникает вполне
целесообразный результат.
Отнюдь не
обязательно, чтобы
коллективное поведение
было наиболее
оптимальным с
нашей точки
зрения. Вспомним,
так ли
уж рационально
устроено все
в человеческом
обществе? И
тем не
менее человечество
живет. И
общественные насекомые
должны вести
себя таким
образом, чтобы
этого было
достаточно для
выживания в
процессе естественного отбора. Не
более того.
Итак,
наука пока
располагает лишь
самыми общими
представлениями о
принципах организации
коллективного поведения
общественных насекомых.
В будущих
исследованиях этой
проблемы специалистам, несомненно,
поможет
теория неравновес-
378
ных процессов, созданная недавно известным бельгийским физиком и математиком И. Пригожиным. Она-то и позволяет понять, как возникает порядок из хаоса. В последние годы достигнуты определенные успехи в компьютерном моделировании поведения муравьев при мобилизации по следу на основе некоторых положений этой теории. Но это еще самые первые шаги в данном направлении. Намеченный путь заманчив, хотя и нелегок. Хочется верить, что он привлечет внимание молодых, влюбленных в науку исследователей, которые смогут пройти его до конца.
НА УРОВНЕ СВЕРХОРГАНИЗМА
Колония муравьев проходит те же стадии, как и жизнь организма: она возникает, развивается и погибает. Ф. Кнауер. Муравьи |
В
1911 г.
вышла из
печати статья
В. Вилера
«Колония муравьев
как организм».
В ней
автор впервые сформулировал и
отстаивал идею,
получившую в дальнейшем
наименование
концепции суперорганизма,
или сверхорганизма.
Согласно этой
концепции, семья,
колония общественных
насекомых представляет собой
настоящий
организм, вполне сопоставимый
по
составу, opганизации
и основным
жизненным функциям
с обыкновенным
организмом животного
или растения.
Действительно,
организм многоклеточного
животного слагается
из клеток,
специализирующихся
на
выполнении определенных
функций и
объединяющихся для
этого в
ткани и
органы. В сверхорганизме —
колонии, где
«клетками» являются
насекомые, мы
также находим
подобную специализацию
— это
разделение на
касты и
функциональные группы.
И в
организме, и
в сверхорганизме
налажена передача
информации между
элементами: нервные
импульсы, гормоны,
вещества-метаболиты —
в организме;
химические, зрительные,
тактильные сигналы
— в
колонии общественных
насекомых. Именно
эти механизмы
коммуникации и
связывают все
части как
организма, так
и сверхорганизма
в единое
целое, позволяют
поддерживать стабильность
их основных
свойств. Обе
системы обладают
выраженной индвидуальностью
и своеобразием
строения. Все
знают, например,
что каждый
из нас
уникальное творение
природы, любой
человек неповторим
и чем-то
отличается от
остальных людей.
Это справедливо
и по
отношению ко
всем живым
существам —
организмам. Но такими
же
свойствами обладает
и колония
общественных насекомых.
Специалистам известно,
что каждая
семья, колония муравьев, пчел,
термитов отличается
от других
379
таких
же по
составу, численности,
строению гнезда,
размерам, особенностям
поведения и даже по
«характеру» насекомых.
Сверхорганизм способен
реагировать на внешние воздействия
вполне целесообразно
и вести
себя как
единое целое,
т. е. так же,
как и
организм.
Сверхорганизм,
подобно организму,
сооружает убежище,
добывает корм,
защищается от
врагов, растет
и, проходя
этапы «детства»
и «юности»,
достигает «зрелости»,
когда становится
возможным его
размножение. При
этом заметим,
что назначение
большинства насекомых
— бесплодных
рабочих, как
и множества
клеток в
организме, заключается
в обеспечении
выращивания максимального
числа плодовитых
самок и
самцов, аналогичных
половым клеткам
организма. Жизнь
рабочих, как
и телесных
клеток, важна
только постольку,
поскольку она способствует
продолжению
рода. При
необходимости рабочих,
как и
клетки, приносят
в жертву и они
гибнут для
достижения этой
цели.
Сверхорганизм, как
и
организм, является
единицей естественного
отбора. Семьи,
колонии, которые
успешнее других
обеспечивают
себя
пищей и
противостоят
неблагоприятным
внешним
воздействиям,
быстрее растут
и
оставляют
больше
потомков.
Так
на основе
отбора семей
и колоний
осуществляется
эволюция
общественных
насекомых.3
Размножается сверхорганизм примерно
так же,
как и
обычный организм.
Ведь закладка
гнезд самками
или брачными
парами у
термитов вполне
аналогична половому размножению многоклеточных
животных: самка-основательница
после
спаривания с
самцом —
это эквивалент
оплодотворенной яйцеклетки.
Многие водные
животные —
морские ежи,
звезды, коралловые
полипы и
даже рыбы
— выпускают
половые клетки
в воду,
где и
происходит оплодотворение,
а затем
из каждой оплодотворенной яйцеклетки
развивается новый
организм. Сходным
образом из
гнезда общественных
насекомых вылетают
самцы и
самки, и
после спаривания
каждая самка основывает свое гнездо.
Да
и развитие
новой колонии
от одной
самки с
постепенным возрастанием
количества насекомых в ней
и все
большим разделением
труда очень напоминает
развитие
нового организма
из яйцеклетки
путем непрерывного
увеличения числа
клеток, возникновения
различий между
ними, формирования
тканей и
органов. Размножение
роением, отделением
отводков, разделением колонии вполне
аналогично бесполому
размножению животных:
почкованию у
гидры и
других полипов,
развитию целого
животного из
части тела
у червей,
морских звезд.
Подобных
аналогий между
сверхорганизмом и
организмом можно
привести много.
С точки
зрения концепции
сверхорганиз-
3 Каждый раз, когда речь идет об эволюции общественных насекомых, мы сознательно упрощаем это явление (см. примечание к первому разделу).
380
ма
колония общественных
насекомых —
это почти
то же
самое, что
и колония
коралловых полипов,
сифонофор или
мшанок. Все
это сверхорганизмы,
состоящие из обычных организмов,
между которыми
происходит разделение
функций и
специализация. Вспомним,
что многоклеточные
организмы возникли
в эволюции
из одноклеточных:
дочерние клетки,
образующиеся при
делении материнской,
остались вместе
и сформировали
единый организм.
Точно так
же возникают
и сверхорганизмы:
потомки самки-основательницы
(или,
например, дочерние
коралловые полипы)
остаются вместе
с ней
в колонии.
Можно представить
себе иерархическую
систему из
трех уровней
организации: одноклеточный
организм —
многоклеточный организм — сверхорганизм.
На каждом
следующем уровне
организмы состоят
из элементов,
которые самостоятельно
существуют на
предшествующем уровне
(организмы первого
уровня — клетки сложены
неживыми элементами
— молекулами).
В
первой половине
XX в.
многие ученые
поддерживали и
развивали концепцию
сверхорганизма и
очень увлекались
поиском все
новых аналогий
между колонией
насекомых и организмом животного
или растения.
Постепенно, однако,
отношение к
этой концепции
начало меняться.
Прежде всего
стала ясной
ошибочность утверждения
Вилера, что
колония животных
именно организм,
а не
просто аналог
индивида. На
самом деле
это системы
двух разных
уровней организации.
Сверхорганизм сам
образован из
организмов и
никак не
может быть
эквивалентом своих
составных частей.
Это противоречило
бы современным
представлениям об
устройстве природы.
Мы всегда
должны помнить,
что за
множеством далеко
идущих аналогий
в организации
и жизнедеятельности
организма и
сверхорганизма
может
скрываться
совершенно
иная суть
явлений и
процессов,
глубоко различных
по
происхождению
и организации.
К
тому же
помимо аналогий
мы при
внимательном
анализе
находим и
множество существеннейших
различий между
сверхорганизмом
и
организмом.
Наиболее важные из
них связаны
с генетикой.
В организме
все клетки
в норме
имеют одинаковый
набор генов,
т. е.
генетически идентичны,
поскольку происходят
от одной
яйцеклетки, многократно
разделившейся бесполым
путем. А
вот в
колонии общественных
насекомых каждый
ее член
генетически уникален,
обладает собственным
генотипом, отличающим
его от
собратьев, поскольку
появляется на
свет в
результате полового
размножения.
При
моногинии
все
рабочие
—
сестры,
а
при полигинии
могут быть
менее родственными
друг другу.
Кстати, колонии
кораллов, мшанок
и сифонофор
в этом
отношении гораздо
ближе к настоящим организмам.
Ведь они
образуются путем
бесполого размножения, и все
их члены
генетически идентичны.
Сверхорганизм
в отличие
от организма
состоит из
особей, по
крайней мере,
двух (а
может быть,
и более)
генетически различ-
381
ных поколений — материнского и дочернего. Рабочие в колонии общественных
насекомых нередко могут оставлять потомство, причем оно появляется на свет
в результате полового процесса и значительно отличается по генетическим свойствам
от потомства царицы. Все это оказывается чрезвычайно важным при анализе происхождения
и эволюции социального поведения насекомых.
Во второй половине XX в. всеобщее увлечение концепцией сверхорганизма сменилось
другой крайностью — ее почти полным забвением. Но перегибы в науке вредны!
И постепенно в научном сообществе восторжествовало более здравое отношение
к концепции. Стало ясно, что во многих случаях она может быть полезной. Например,
при изучении экологии общественных насекомых и их связей с другими компонентами
экосистем удобно рассматривать колонии как сверхорганизмы. При этом в экологических
взаимоотношениях с прочими животными и растениями они во многом эквивалентны
обычным организмам. Мы можем легко представить себе семью медоносных пчел,
ос, колонию муравьев как своеобразный организм, обитающий в построенном им
гнезде, но добывающий корм на большой территории вокруг, посылая за ним составляющих
его насекомых. Этакое огромное животное, некое подобие морской лилии или
актинии, вылавливающей из окружающей воды добычу щупальцами, либо одноклеточной
корненожки, выпускающей во все стороны ложные ножки — псевдоподии! При исследовании
потребления питательных веществ, процессов почвообразования, участия в пищевых
цепях и круговороте вещества понятие о колониях общественных насекомых как
о сверхорганизмах очень продуктивно.
Концепция сверхорганизма может пригодиться и при изучении развития насекомых
разных каст, разделения функций между ними, сохранения целостности и единства
колонии. Ведь все эти процессы во многом сходны с тем, что мы наблюдаем в
обычном организме. Как и организм, представляющий собой нечто большее, чем
сумму клеток, колония не просто совокупность насекомых. Ее члены связаны
многочисленными взаимодействиями, зависят друг от друга, и это придает колонии
как организованному целому совершенно новые свойства, которыми не обладают
составляющие ее насекомые. Мы уже рассматривали примеры возникновения новых
свойств при объединении насекомых в «Феномене коллектива», обсуждали способы
коммуникации, пути распределения пищи, механизмы регуляции развития и другие
формы взаимозависимости общественных насекомых. Теперь же остановимся на
так называемом эффекте группы, т. е. влиянии присутствия особей своего вида
на физиологическое состояние и поведение отдельного насекомого.
Оказывается, подобные взаимодействия нередко значительны. Многие формы поведения
могут проявляться только при
382
определенной минимальной численности группы насекомых. Так, для того чтобы
пчелы начали сооружать сот, их должно быть не менее 50, если среди них
есть матка, а при ее отсутствии строительство развернется только при
наличии 10
тыс. рабочих. Образование роя возможно, когда пчел не менее 200. Изолированное
общественное насекомое зачастую ведет себя совершенно иначе, чем в группе.
Пчелы и муравьи,
например, в одиночестве становятся неагрессивными и даже не реагируют
на феромоны тревоги. Для проявления нормальных оборонительных реакций
тоже
необходима минимальная численность группы. Недаром О. Форель еще в 1874
г. писал: «Всякий
обитатель большого муравейника значительно смелее своего такого же собрата
из маленького муравейника. Всякий муравей пойдет десять раз на смерть,
если он окружен товарищами, и, напротив, будет выказывать чрезвычайную
робость,
избегая малейшей опасности даже перед самым слабым противником, если
он находится в каких-нибудь тридцати шагах от своего гнезда»4.
Объединение насекомых в
группу стимулирует многие физиологические процессы. У рабочих муравьев
и шмелей, если их содержать хотя бы небольшими группами, в отсутствие
подавляющего
влияния царицы яичники развиваются значительно скорее, чем у изолированных
насекомых, к тому же во втором случае это происходит лишь у немногих
особей. А находящиеся в «одиночном заключении» медоносные пчелы вообще
не откладывают
яиц, хотя обеспечены всем необходимым. Самки-основательницы черного садового
муравья начинают откладывать яйца на третий-четвертый день, если они
живут поодиночке, и в первый же день при содержании группами по две,
три или
пять.
Самки в группах быстрее, чем одиночки, выращивают первых рабочих и меньше
при этом теряют в весе. Но при объединении основательниц возникают и
отрицательные эффекты: каждая самка в группе откладывает меньше яиц и
среди них выше
доля кормовых. Хотя это, вероятно, связано не с эффектом группы, а с
иерархией доминирования между самками и подавлением плодовитости низкоранговых
членов
группы.
Эффект группы проявляется и в социальных взаимоотношениях. При увеличении
числа муравьев, термитов, пчел в группе они чаще вступают в трофаллаксис,
кормят личинок, усерднее занимаются строительством и другой деятельностью.
Еще в начале XX в. русский зоолог В. Караваев установил, что муравьи,
когда их не более 10 в формикарии, слабо реагируют на освещение, тогда
как в
группах с численностью 50—100 они тут же приступают к эвакуации личинок
из освещенной
камеры в темную. В 1937 г. китайский энтомолог Чи Занчен обнаружил эффект
группы при строительстве у японского кампонотуса. Посаженные по одному
в баночки с песком рабочие «раскачивались» два-три часа и за шесть часов
выкапывали
очень немного
4 Forel A. Les fourmis de la Suisse. Zurich, 1874. P. 381.
383
песка. Но в группах по двое и по трое они принимались за дело уже через
полчаса и иногда увеличивали производительность труда более чем
в 3000 раз! Кстати,
среди муравьев были и ретивые землекопы-«трудяги», и «лодыри».
Так вот, если их сажали вместе, то «трудягам» в некоторой степени удавалось
«перевоспитать»
ленивых муравьев и те тоже брались рыть песок.
В дальнейшем было установлено, что не всегда производительность
труда при группировке повышается. Так случается, как правило, только
в
группах старых
рабочих. Молодые внутригнездовые рабочие склонны собираться по
нескольку вместе и столь долго заниматься взаимным туалетом, что
для работы
времени почти не остается. Кроме того, производительность труда
зависит от
размера садка и числа муравьев: в небольшом сосуде или же когда
рабочих слишком
много, они тратят много времени на контакты друг с другом, а в
процессе работы мешают
своим товарищам, что снижает эффективность их совместной деятельности.
Эффект группы зачастую обусловлен простым присутствием других насекомых
своего вида, и для его проявления необязательны контакты между
членами группы. Вероятно,
насекомые узнают о наличии собратьев в основном по запаху. Когда
муравьев формика йессензис разделили непрозрачной сеткой, они продолжали
стимулировать
друг друга к рытью почвы, а разграниченные стеклом вели себя так,
будто находятся в одиночестве. Возможно, при рытье из-за движений
челюстей
мандибулярные железы выделяют свой секрет, он и оказывает на муравьев
группы стимулирующее
действие. В экспериментах с рабочими лазиуса они, отделенные от
большой группы
муравьев своего гнезда сеткой, значительно чаще кормили личинок,
чем оказавшиеся за стеклом.
В опытах автора с самками-основательницами черного садового муравья
им предоставляли различные «жилищно-бытовые» условия. Каждая самка
в первом
варианте получала
«отдельную квартиру» — прикрытую стеклом гипсовую камеру 10X15
мм, высотой 10 мм. Во втором варианте самки жили в «коммунальных
квартирах»
— камерах
20X30 мм той же высоты, разделенных пластиковыми перегородками
на четыре одинаковые «комнаты», в каждой из которых обитала одна
«квартиросъемщица».
Площадь и объем «комнат» были точно такими же, как и «отдельных
квартир». Но жильцы «коммуналок» могли ощущать присутствие соседей
по запаху,
проникавшему сквозь тонкие щели над и под перегородками, а также,
возможно,
по сотрясению
перегородок. Оказалось, что эффект группы полностью обнаружился
в «коммунальных квартирах». Там быстрее, чем в «отдельных квартирах»,
появились первые
яйца и их в сумме было гораздо больше, а за 40 дней опыта не погибла
ни одна самка.
Среди обитательниц «отдельных квартир» 40% до конца эксперимента
не отложило яиц и половина из них умерла.
384
Значит, для возникновения эффекта группы и в этом случае нет необходимости
в контактах между самками, достаточно лишь восприятия присутствия
других особей на расстоянии.
Эффект группы ярче всего проявляется, конечно, у наиболее высокоорганизованных
эусоциальных видов. В изоляции от собратьев муравей, пчела
или термит не только ведет себя совершенно иначе, но и нередко
не
выживает. Взаимозависимость отдельных насекомых, составляющих
сверхорганизм,
у этих видов столь
же велика, как и клеток обычного организма.
Колония общественных насекомых, как и организм, является системой,
перерабатывающей добываемую во внешней среде пищу в те специфические
вещества, из которых
построена сама, чтобы обеспечить свой рост, достичь необходимой
для размножения величины и оставить возможно больше плодовитых
потомков.
Естественный
отбор, которому подвергаются семьи и колонии общественных насекомых,
способствует
успешному решению этой задачи, совершенствуя стратегии роста
и размножения
у разных видов.
Для галиктов, шмелей, очень многих ос и других насекомых с
ограниченным циклом развития колонии характерен взрывной
тип роста численности.
При этом сначала
быстро нарастает количество рабочих, затем колония переключается
на выращивание плодовитых насекомых, затрачивая на них все
доступные ей
ресурсы, и наконец
распадается по мере гибели рабочих. При осциллирующем росте вначале тоже быстро увеличивается количество рабочих, а затем
колония
вступает в стационарное
состояние, т. е. численность рабочих подвержена довольно правильным
циклическим колебаниям, которые могут быть вызваны двумя причинами.
У большинства
муравьев и термитов число рабочих немного уменьшается в период
выращивания крылатых,
поскольку оно поглощает значительную часть ресурсов колонии
и на появление новых рабочих их остается мало. После вылета
самцов
и
самок количество
рабочих постепенно увеличивается вплоть до нового сезона размножения.
У медоносных
и безжалых пчел, многих ос, размножающихся роением, у муравьев
и термитов, образующих новые гнезда путем отделения отводков
или делением
материнской
колонии примерно пополам, колебания численности вызваны потерей
части рабочих (иногда свыше 50%), уходящих в отводок или улетающих
с роем.
Для того чтобы число рабочих увеличивалось, вновь появляющихся
насекомых должно быть больше, чем погибших за то же время.
Продолжительность жизни и смертность рабочих неодинаковы у
разных видов. Рабочие
обыкновенной
осы в среднем живут около девяти дней, а смертность среди них
достигает 10%
в сутки. Количество рабочих в гнезде может к концу лета превысить
2 тыс.,
но
для этого осы выращивают в сумме более 10 тыс. рабочих. У лесной
осы смертность рабочих меньше и они живут в среднем 14 дней,
однако цикл
развития вида
385
значительно короче, чем у обыкновенной осы. Поэтому максимальное
число рабочих в гнезде составляет 150—200, а всего их выращивают
300—350.
Средняя продолжительность
жизни рабочих полевого шмеля около 25 дней при смертности
4% в сутки. Смертность медоносных пчел летом примерно 3% в день.
В колонии
фараонова
муравья ежедневно
погибает около 2% рабочих, а у муравья-кочевника эцитон
хаматум приблизительно 1 %.
Что определяет максимально возможную величину колонии общественных
насекомых? Прежде всего, конечно, продуктивность царицы.
Чем больше она откладывает
яиц, тем больше новых рабочих может быть в гнезде. Поэтому
при развитии колонии она сначала растет быстро, но со временем
рост
постепенно
замедляется и совсем
прекращается, когда количество появляющихся за определенный
промежуток рабочих уравнивается с числом погибающих. Продуктивность
царицы,
как правило, сильно
повышается при увеличении количества рабочих в гнезде,
что задерживает остановку роста колонии и позволяет ей
достигнуть
предельных
размеров. Следует учитывать,
что гибнут не только взрослые насекомые, но и яйца, личинки,
куколки. У полевого шмеля, например, личинки появляются
из 71% яиц, 73%
личинок доживает
до стадии
куколки и из 90% куколок выходят рабочие, т. е. лишь 47%
яиц развивается во взрослых насекомых. Для рыжей мирмики
эти цифры
соответственно
50, 74 и 89%, т. е. у нее рабочие муравьи развиваются в
среднем из 33%
отложенных яиц. Гибнут личинки чаще всего при очередной
линьке.
Высокая смертность части расплода связана, конечно, с пониженной
жизнеспособностью отдельных яиц, личинок, куколок, но,
кроме того, зависит от обеспеченности
колонии пищей. Царицы многих видов откладывают больше яиц,
чем можно вырастить личинок при наличных ресурсах. Когда
не хватает
провизии,
рабочие-няньки уничтожают часть яиц или личинок. При этом
они оставляют личинок покрупнее,
на выкармливание которых нужно уже немного пищи, и поедают
мелких и слабых, выделяющих, вероятно, меньше специфических
феромонов.
Муравьи в случае
возникновения «продовольственной проблемы», как уже говорилось,
нередко просто не кормят
мелких личинок, а те не растут, ожидая лучших времен, или
же впадают в
диапаузу и остаются зимовать, чтобы продолжить развитие
следующим летом.
Все это позволяет
«подгонять» количество выращиваемых личинок под доступные
пищевые ресурсы и увеличивать скорость «оборота капитала»,
т. е. вкладывать
усилия
и корм таким образом, чтобы скорее получать новых рабочих,
которые сами
станут
выращивать личинок и добывать пищу.
Итак, максимально возможный размер колонии зависит от продуктивности
царицы, выживаемости расплода и взрослых насекомых. Эту
предельную величину можно
повысить, напри-
386
мер, совершенствуя стратегию добывания корма, используя
более эффективные формы мобилизации, коллективную охоту,
вторичное
деление территории
и т. д. Можно улучшить и эффективность выращивания
личинок. Наибольших результатов
в этом направлении добились, вероятно, медоносные пчелы,
у которых из
85% отложенных яиц развиваются взрослые рабочие. Наконец,
третий путь заключается
в повышении продуктивности единственной царицы или
увеличении их числа. Полигиния в принципе позволяет колонии достичь
огромных размеров. Но
по мере возрастания
численности рабочих уменьшается их эффективность, во-первых,
потому,
что скученность приводит к столкновениям и взаимным
помехам в деятельности, а во-вторых, из-за того, что рабочие
вынуждены отправляться все
дальше от гнезда
за пищей для приумножающегося населения. Одним выходом
из этого положения является уменьшение численности
насекомых в колонии
путем роения,
отделения отводков, а вторым — возникновение полидомности,
федераций
и других
развитых форм социальной структуры, которые мы уже
рассматривали.
Переход к размножению, т. е. созревание колонии общественных
насекомых, совпадает с тем моментом, когда численность
рабочих достигает такой
величины, что царица
или царицы уже не в силах эффективно сдерживать выращивание
плодовитых самок и подавлять плодовитость всех рабочих.
В это время многие
рабочие, правда,
далеко не у всех видов, откладывают яйца, из которых
появляются самцы.
Рост и размножение организма и колонии общественных
насекомых, или сверхорганизма, как мы видим, могут
быть описаны
в сходных понятиях.
И тот и другая являются
объектами изучения общей теории систем. В последние
10—15 лет достигнуты серьезные успехи в математическом
и компьютерном
моделировании процессов роста, развития и размножения
колоний ос, шмелей, пчел,
муравьев.
Очень интересно также сравнение способностей организма
и сверхорганизма к поддержанию своей устойчивости.
Известно, что в организме
существуют сложные
физиологические механизмы, направленные на стабилизацию
химического
состава крови и лимфы, кровяного давления, температуры
тела и других параметров
внутренней среды. Процесс поддержания постоянства всех
этих параметров в организме называется
гомеостазом. У общественных насекомых есть аналогичное
явление — социальный гомеостаз. Он выражается
прежде всего в поддержании
неизменных
численных
соотношений между насекомыми разных каст, между взрослыми
рабочими и личинками. Конечно, это возможно в стабильных
условиях, когда
колония достигла стационарного
состояния. Мы уже говорили о том, что доля солдат среди
всех насекомых бесплодной касты у термитов и муравьев
величина довольно постоянная,
и обсуждали механизмы,
387
которые ее регулируют. В природных колониях рыжей мирмики
весной отношение количества рабочих к числу личинок
в среднем составляет
0,88. Лабораторные
эксперименты показали, что эффективность выкармливания
личинок максимальна, когда на 50 личинок приходится
42 рабочих, т.
е. при соотношении
рабочих и личинок 0,84, которое очень близко к
величине, наблюдающейся в природе.
Значит, в естественных условиях производительность
труда рабочих близка к максимальной, а гомеостатические
механизмы
(какие
— мы пока не знаем)
поддерживают
оптимальные соотношения между численностью рабочих
и личинок.
У моногинных видов существуют те или иные способы
сохранения моногинности: агрессивность царицы или
рабочих к лишним
плодовитым самкам, феромоны,
препятствующие развитию новых цариц, роение или
почкование и т. д. А для полигинных колоний
характерны определенные численные соотношения между
царицами и рабочими. В частности, у рыжей мирмики
на одну царицу
приходится в среднем
76 рабочих.
Если кормить термитов пищей с малым содержанием
азота, то они прибегнут к каннибализму и будут
поедать часть
личинок и взрослых
собратьев,
в тканях которых азота достаточно. Конечно, это
временная мера, которая дает термитам
возможность продержаться без поступления азота
извне. Точно так же при
голодании
муравьи поедают яйца, личинок и даже куколок. Эти
гомеостатические механизмы сверхорганизма напоминают
то, что происходит
в
обычном организме. Например,
при недостатке пищи используются резервные жировые
запасы, а потом и другие ткани; если не хватает
кальция, организм
добывает его
из собственных костей.
Многие термиты съедают оставшихся крылатых после
окончания благоприятного
для вылета сезона. То же самое наблюдается у животных,
когда рассасываются их половые клетки, не использованные
во время
брачного периода.
Можно привести и другие примеры социального гомеостаза.
Пожалуй, больше всего мы знаем о регулировании
микроклимата и иных
параметров среды
в гнездах общественных
насекомых.
Самые примитивные общественные насекомые неспособны
активно изменять физические условия в гнездах и
полностью зависят
от внешней среды.
В жилище пчел-галиктов
температура и влажность воздуха такие же, как и
в норке, вырытой поблизости одиночной пчелой. Единственное,
что могут делать
галикты, это выбирать
для гнездования участки, которые достаточно, но
не
чрезмерно прогреваются солнцем
и имеют подходящую влажность почвы (только некоторые
виды умеют вентилировать свои норки крыльями).
По сути,
так
же поступают
аллодапиновые пчелы,
стеногастрины и другие примитивные осы и даже муравьи
и термиты некрупных семей, сооружающие
подземные гнезда. Но большинство видов общественных
насекомых способно в той или иной мере регулировать
микроклимат
своих
388
жилищ и в первую очередь температуру. Для этого
они используют два принципа. Долговременная
регуляция основана на устройстве
самого гнезда, его теплоизолирующих
свойствах, возможностях вентиляции и, конечно,
на
выборе для строительства наиболее подходящего
участка. Кратковременные
регулирующие механизмы реализуются через поведение насекомых, способных
либо переносить расплод в оптимальные
в данный момент по микроклимату зоны гнезда,
либо активно изменять
в нем температуру, влажность и газовый состав
воздуха.
Осы-полисты, несмотря на невысокую численность
рабочих, довольно эффективно регулируют температуру.
Они строят
гнезда в хорошо
прогреваемых местах,
поскольку для развития их личинок благоприятна
температура около 34—35°С. Но в жаркий
летний день сот может перегреться. Как только
его температура достигает 35°С, осы начинают
быстро
махать крыльями,
обдувая и охлаждая сот
воздухом. Чем
больше приток тепла к гнезду, тем интенсивнее
эта вентиляция. Но при сильном перегреве ее
уже недостаточно.
И тогда
осы приносят воду и
распределяют капельки по соту. Вода при активном
обдувании сота вмиг испаряется и
охлаждает
его.
В результате осам удается в течение всего солнечного
дня поддерживать на соте температуру не ниже
34 и не выше 37,5°С,
что позволяет
расплоду успешно
развиваться. Разумеется, ночью и в прохладные
дни температура гнезда
опускается значительно ниже оптимума, ведь
осы не могут его согревать. Интересно,
что во время дождя, когда на сот попадает вода,
полисты резко изменяют поведение.
Только что они приносили в гнездо воду, чтобы
защитить его от полуденной жары. Но вот грянул
гром и забарабанили
первые
капли
грозового
ливня. И в жилище полистов «объявляется аврал»,
ведь дождь может намочить
осиную бумагу
и гнездо разрушится. Поэтому все осы дружно
борются с «наводнением». Обнаружив
каплю воды на соте, оса тут же выпивает ее,
а затем, свесившись вниз с сота, выплевывает ее
из зобика.
Только так им удается
спасти свое
жилище. Некоторые
полибии в дождь сидят на открытом соте, свесив
брюшки вниз, и вода стекает по их телам и крыльям.
У ос, строящих гнезда с теплоизолирующей оболочкой,
терморегуляция гораздо совершеннее. Эти осы
умеют не только охлаждать
жилище посредством вентиляции
и разбрызгивания воды по сотам, но и согревать
его теплом собственных тел. Любое насекомое
в процессе жизнедеятельности
выделяет
тепло (оно называется
метаболическим), но его так мало, что насекомые
являются
холоднокровными животными. Интенсивное сокращение
мышц дает значительно больше
тепла. Поэтому в полете многие насекомые благодаря
работе мощной крыловой
мускулатуры становятся по существу теплокровными.
Например, температура груди летящего
шмеля достигает
35—37°С даже при температуре воздуха всего
10—15°С. Общественные насе-
389
комые научились коллективно использовать метаболическое тепло для обогрева
гнезда, и многие высокоорганизованные их виды на уровне сверхорганизма
являются по-настоящему «теплокровными». Это позволяет им содержать расплод
в стабильных
условиях, что увеличивает выживаемость потомства. Кроме того, повышенная
температура ускоряет развитие яиц, личинок, куколок, а следовательно, и
рост всей колонии. Но «теплокровность» имеет место только в гнезде. Покидая
его,
насекомое вновь становится холоднокровным (это не относится к шмелям).
«Теплокровность» семьи обыкновенной осы еще не совсем совершенна. Температура
в жилище этих насекомых в некоторой степени зависит от погоды. Когда наружная
температура опускается до 9—10°С, в гнезде она остается выше 26°С. В жаркую
погоду соты никогда не перегреваются выше 34°С даже при внешней температуре
воздуха до 40°С. Среднесуточная температура окружающего гнездо воздуха
в течение всего северного лета не превышает 18—20°С, в то время как на
сотах
она в среднем составляет 30—32°С, обеспечивая оптимальные условия для развития
расплода.
Примечательно, что даже самка-основательница веспин способна согревать
гнездо вскоре после его постройки. Свернувшись «калачиком» вокруг стебелька
сота
под защитой оболочки и попеременно втягивая и растягивая брюшко для усиления
дыхания, она выделяет тепло и повышает температуру в гнезде в среднем на
1°С по сравнению с окружающей. Когда появляются личинки, благодаря их метаболическому
теплу температура в гнезде становится уже на 4°С выше.
Шмели и многие безжалые пчелы тоже используют метаболическое тепло для
согревания гнезд, а вентиляцию крыльями — для охлаждения. Но понижать температуру
путем
испарения воды они не умеют. Безжалые пчелы, живущие некрупными семьями,
не могут регулировать микроклимат и довольствуются тем, что выбирают для
поселения наиболее подходящие места. Другие же виды, гнезда которых окружены
толстой оболочкой или иными теплоизолирующими материалами, способны поддерживать
в них температуру 33—36°С даже при похолоданиях до 8—10°С. Это позволило
отдельным из них заселить в Южной Америке районы с морозными зимами. В
жаркие дни пчелы усиленно вентилируют жилище крыльями, создавая громкий
шум. Иногда
они даже проделывают в оболочке гнезда дополнительное отверстие напротив
входа, чтобы обеспечить воздухообмен.
Приуроченные в основном к странам с умеренным и даже холодным климатом
шмели прежде всего, конечно, сталкиваются с необходимостью обогрева гнезд.
Этому
способствует гнездование в почве, дуплах и использование значительных количеств
утепляющих материалов. Рабочие шмели обогревают личиночники и коконы, плотно
прижимаясь к ним всем телом и совершая быстрые дыхательные движения брюшком.
Даже одиноч-
390
ная самка-основательница может путем такого «насиживания» повысить температуру
личиночника более чем на 5°С. На личиночнике постепенно образуется
канавка, в которую плотно входит тело самки. В крупном же гнезде, где рабочих
много, терморегуляция приближается к почти идеальной «теплокровности».
Например,
при наблюдениях за большой семьей полевого шмеля было установлено,
что
температура в ее жилище поддерживалась на уровне 30°С с колебаниями
в обе стороны на
полградуса и очень мало зависела от наружной температуры. Открытые
соты гигантской и карликовой пчел почти постоянно покрыты плотным «одеялом»
из двух-трех,
а иногда и более слоев рабочих, защищающих расплод от солнечных лучей,
капель дождя и способствующих сохранению тепла. В жаркое время пчелы
охлаждают сот,
вибрируя крыльями и разбрызгивая по нему воду, а при похолодании сбиваются
в кучу и усиливают выделение тепла. Это позволяет им поддерживать на
соте температуру 30—31°С у гигантской и 33—37°С у карликовой
пчелы.
Наиболее совершенной терморегуляцией среди летающих общественных насекомых
обладают, конечно, обыкновенная и индийская медоносные пчелы. С ранней
весны до осени они поддерживают на сотах температуру от 34,5 до 35,5°С,
как в очень
точном термостате! Каждая пчела выделяет в среднем около 0,4 Дж метаболического
тепла в минуту, а вся семья — больше 10 кДж. Этого вполне достаточно
в теплую погоду. При похолоданиях пчелы усиливают теплопродукцию, вибрируя
крыловой
мускулатурой. В это время, приложив ухо к улью, можно услышать равномерное
и довольно громкое гудение. Сохранению тепла в гнезде способствует
его конструкция: дупло обладает хорошими теплоизолирующими свойствами,
а
пчелы
тщательно заклеивают
прополисом все трещины и щели, сужают вход, сводя потери тепла к минимуму.
Зимой пчелы сбиваются в шарообразный клубок, что позволяет им расходовать
еще меньше тепла и поддерживать в центре клуба температуру выше 20°С.
Пчелы почти постоянно вентилируют свое жилище. Возле летка и снаружи,
и внутри гнезда стоят пчелы, быстро машущие крыльями. У одного края
летка они обычно
обращены головами к нему, а у другого — от него. Поэтому возникает
правильный ток воздуха: он входит в леток, омывает соты и выходит с
противоположной
стороны летка. Поддержанию этого круговорота способствуют и вентилирующие
пчелы на сотах. Такое проветривание нужно не только для охлаждения
улья, но и для удаления образующейся там углекислоты и пополнения запасов
кислорода.
Кроме того, при вентиляции улей освобождается от воды, испаряющейся
при приготовлении меда из нектара.
Когда становится слишком жарко, пчелы, продолжая вентилировать свое
жилище, начинают распределять по сотам капель-
391
|
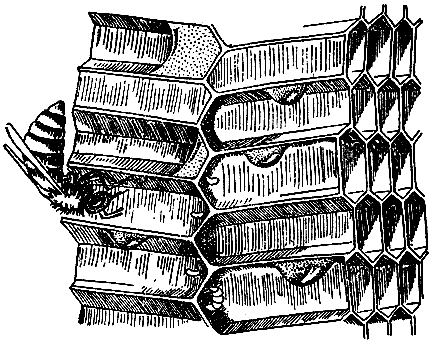
ки воды. Обычно они подвешивают их внутри пустых ячеек, а также
ячеек с яйцами или маленькими личинками, реже капли помещают между
крышечками
запечатанных
ячеек. До тех пор пока пчелы имеют свободный доступ к воде, они
способны поддерживать в улье нормальную температуру, несмотря на
любую жару.
В опыте, проведенном на юге Италии, улей с пчелами поставили в
полдень на раскаленную
солнцем бетонированную площадку, температура поверхности которой
достигала 70°С. И что же? Температура в улье ничуть не изменилась
и в течение
нескольких
часов, пока длился опыт, оставалась 35°С! Вереницы пчел-водоносов
летали при этом из улья к находящемуся неподалеку фонтану, и доставляемая
ими вода
позволила семье пчел выдержать сверхэкстремальные условия.
Муравьи и термиты лишены возможности вентилировать жилища крыльями,
но метаболическое тепло выделяют, как и все животные. Терморегуляция
в их
гнездах осуществляется
иначе, чем у пчел и ос. Большинство видов муравьев умело использует
для этих целей градиент температуры, создающийся в их гнездовых
холмиках, или помещает
расплод под камнями, играющими роль аккумуляторов солнечного тепла.
Только
благодаря этому муравьи способны процветать в странах с умеренным
климатом, хотя, как правило, для их нормального развития требуется
температура
не ниже 25°С. Некоторые муравьи смогли проникнуть даже в районы
Крайнего Севера,
где всегда поселяются на хорошо прогреваемых солнцем участках и
строят гнезда
так, чтобы максимально использовать достигающую их поверхности
солнечную радиацию. А вот в густых лесах умеренной зоны и горных
тропических
лесах очень мало видов муравьев, и они живут там всегда на прогалинах
или полянах,
согреваемых солнечными лучами.
Лишь немногие виды муравьев достигают такой численности рабочих
в колонии, что способны заметно повышать температуру в гнезде.
Наилучшим
образом
исследована эта терморегуляция у рыжих лесных муравьев. Во внутреннем
конусе их муравейника
с ранней весны до конца лета существует так называемое тепловое
ядро — область повышенной температуры, где находится основная масса
куколок
и
личинок. Температура
теплового ядра слабо изменяется в связи с колебаниями внешней
392
температуры. Самая высокая температура, до 30—32°С, бывает в теплые
ночи, когда в муравейнике собираются почти все фуражиры и выделение
метаболического
тепла увеличивзется. Ниже 25—26°С температура в тепловом ядре
не опускается даже при очень холодной погоде.
Долгое время среди специалистов шли споры о том, каким образом
рыжие лесные муравьи поддерживают температуру в своих гнездах.
Постепенно
выяснилось, что терморегуляция у них — очень сложное, комплексное
явление, включающее
множество факторов. Муравейник рыжих лесных муравьев обладает
плотной оболочкой
с хорошими теплоизоляционными свойствами. Муравьи никогда не
поселяются в сильно затененных местах, и солнечный свет, издающий
на купол,
приносит довольно
много тепла. Недаром муравейники всегда располагаются с южной
стороны деревьев, а их более длинный скат тоже обращен на юг!
Значительное
количество тепла
выделяют также микроорганизмы, размножающиеся в перегнивающем
растительном материале, из которого состоит купол муравейника.
По оценкам специалистов,
микроорганизмы дают иногда до 1/2 и даже 2/3 тепла, необходимого
для поддержания температуры в гнезде.
Но все-таки выделение тепла самими муравьями играет наибольшую
роль. Это становится очевидным, если проследить, как сделал
Г. М. Длусский,
за разогреванием
муравейников ранней весной. Как только стает снег на куполе,
масса муравьев высыпает туда и «нежится» на весеннем солнце.
Раньше думали,
что, нагревшись,
муравьи уходят в муравейник и, остывая, согревают гнездо. Их
даже назвали теплоносцами. Простейшие подсчеты показывают,
однако, что
таким путем
много тепла не перенесешь и муравейник существенно не обогреешь.
Значит, «солнечные
ванны» муравьи принимают с какой-то другой целью.
Разогревание муравейника происходит в течение нескольких часов,
причем нередко ночью или в плохую погоду, когда солнца нет.
Температура в центре купола
быстро повышается от 10—12 до 25—30°С. Ясно, что это возможно
лишь за счет выделения муравьями метаболического тепла. Недаром
разогреваться
весной
способны только достаточно крупные муравейники, в которых много
муравьев
с обширными
запасами сладкой пади в зобике и жировых клеток в брюшке, накопленными
еще прошлым летом. Именно сахара в первую очередь «сгорают»
в организме насекомых,
согревая муравейник. После исчерпания их в ход идут жиры. Маленькие
или
малонаселенные муравейники не могут разогреться ранней весной.
Когда температура в гнезде
повысится, начинают размножаться и микроорганизмы, выделяя
тепло «в помощь» муравьям. Способность к разогреву гнезд и
поддержанию
высокой
и довольно
стабильной температуры в тепловом ядре позволяет рыжим лесным
муравьям приступать к зктивной жизнедеятельности и выращиванию
рас-
393
плода уже ранней весной, когда все остальные виды муравьев
еще находятся в неактивном состоянии, и продолжать его
до конца лета.
Однако ведь муравейник может и перегреваться в жаркую погоду
и особенно на солнце. Тогда муравьи усиливают вентиляцию,
открывая на куполе
новые отверстия.
Зарывая в купол муравейника слабые электрические нагреватели,
немецкий
исследователь К. Хорстманн с удивлением обнаружил, что
муравьи обладают неожиданно высокими
способностями к охлаждению гнезда. Муравейники, лишенные
их обитателей, перегреваются уже при мощности нагревателя
2—5
Вт, а населенные
муравьями сохраняют нормальную
температуру даже при подогреве изнутри мощностью 20 Вт.
Как же муравьям удается избавляться от излишнего тепла? Оказалось,
что
муравьи эвакуируют
из купола
вглубь расплод, освобождая все пустоты, и это значительно
улучшает вентиляцию. Кроме того, рабочие, испаряя при дыхании
много
воды, активно охлаждают
муравейник.
Согревание жилища метаболическим теплом возможно и у муравьев
других видов, сооружающих гнезда с хорошими теплоизолирующими
свойствами
и имеющих достаточную
численность рабочих в колонии. Так обстоит дело, например,
у строящих картонные гнезда пахучего муравья-древоточца
и желтого пахучего
муравья. А вот у
муравьев-кочевников терморегуляция осуществляется и без
гнезда. Измерения температуры внутри
скопления муравьев, где находились расплод и царица, на
бивуаке эцитонов показали, что она практически постоянна
и сохраняется
на уровне 29°С,
хотя температура воздуха в этот период колебалась от 24
до 25,5°С.
Конечно, муравьям-кочевникам проще поддерживать необходимую
температуру, ведь
им
«помогает» тропический
климат!
Гнездовые холмики термитов во многих случаях устроены так,
чтобы улавливать и сохранять солнечное тепло, а выделение
метаболического тепла обитателями
термитников не играет существенной роли в терморегуляции.
Таково, например, положение дел у большого закаспийского
термита. Термитник
амитермес
меридионалис имеет форму эллипса, длинная ось которого
направлена с севера на юг,
что позволяет достичь равномерного поглощения тепла в течение
всего дня. Кстати,
примерно такие же форма и ориентация гнездовых холмиков
у желтого земляного муравья в Европе и огненного муравья
в
США. Каким
образом
термиты и
муравьи сооружают гнездо нужной конфигурации и правильно
ориентируют его по сторонам
света, совершенно неясно.
Среди тропических термитов возможности терморегуляции существенно
различаются. Виды, сооружающие небольшие термитники с тонкими
внешними стенками
(роды амитермес, микроцеротермес), неспособны поддерживать
в них постоянную тем-
394
пературу — она практически равна внешней и испытывает такие
же суточные колебания. Напротив, виды, обитающие в
крупных термитниках
с толстыми,
хорошо изолирующими
от наружного воздуха стенками, обладают весьма совершенной
терморегуляцией. Например, в термитниках рода цефалотермес
температура неизменна
и составляет около 29°С, в то время как внешняя температура
колеблется от 22 до
26°С. В обитаемых холмиках термитов рода назутитермес
температура почти на
10°С выше, чем в «мертвых» термитниках. В гнездах австралийских
термитов
рода
коптотермес, расположенных в центре древесных стволов,
температура колеблется от 33 до 38°С, что на 13—20°
выше, чем в таких
же деревьях, но без термитов.
Особенно удивительная система «кондиционирования воздуха»
существует у термитов рода макротермес. В их огромных
термитниках тепло
выделяют как
грибные сады,
так и сами термиты. Поэтому температура в центре гнезда,
где больше всего термитов, поддерживается на уровне
30°С. Теплый,
обогащенный
углекислотой
воздух поднимается в полости над садами и «башни»,
где начинает охлаждаться. Затем он поступает в тончайшие
каналы, вплотную
подводящие к поверхности
термитника, и поры наружных стенок и по ним медленно
опускается к основанию гнезда. В это время воздух дополнительно
охлаждается
до
температуры
24°С и путем диффузии сквозь стенки отдает в атмосферу
углекислоту и обогащается
кислородом. Более прохладный и богатый кислородом воздух
попадает в нижнюю часть термитника — так называемый
погреб,
а из него
опять поднимается
к грибным садам и основной массе обитателей гнезда.
Этот круговорот позволяет
термитам
не только поддерживать необходимую температуру в гнезде,
но и эффективно обновлять его атмосферу. На уровне
земли термиты
проделывают
в
оболочке гнезда отверстие. Если его закрыть, стенки
термитника станут очень
влажными и он
может разрушиться. Вероятно, сухие послеполуденные
ветры способствуют удалению
излишней влаги сквозь отверстие и одновременно охлаждению
термитника. О такой роли этого отверстия свидетельствуют
кристаллы солей,
обычно обнаруживаемые
в погребах и скорее всего выпадающие из почвенных растворов
при испарении воды.
395
СОДЕРЖАНИЕ
| Предисловие ...................................................................................................................................... |
3 |
| ПОРАЗИТЕЛЬНЫЙ МИР ................................................................................................................. |
5 |
| Первое знакомство ............................................................................................................................ |
7 |
| Путь к социальности ......................................................................................................................... |
16 |
| БИОЛОГИЯ И ОБРАЗ ЖИЗНИ ....................................................................................................... |
29 |
| Общественные осы ........................................................................................................................... |
31 |
| Общественные пчелы ....................................................................................................................... |
47 |
| Пчелы-галикты .................................................................................................................................. |
49 |
| Аллодапиновые пчелы ..................................................................................................................... |
54 |
| Шмели ................................................................................................................................................. |
57 |
| Безжалые пчелы ................................................................................................................................ |
66 |
| Медоносные пчелы ........................................................................................................................... |
75 |
| Муравьи .............................................................................................................................................. |
86 |
| Видовое разнообразие и социальная организация ........................................................................ |
89 |
| Муравьиные жилища ........................................................................................................................ |
102 |
| Питание .............................................................................................................................................. |
116 |
| Термиты .............................................................................................................................................. |
128 |
| Видовое разнообразие и социальная организация ........................................................................ |
130 |
| Жилища термитов .............................................................................................................................. |
135 |
| Питание ............................................................................................................................................... |
140 |
| Основание новых гнезд .................................................................................................................... |
144 |
| ОСНОВНЫЕ ПРИНЦИПЫ СОЦИАЛЬНОЙ ОРГАНИЗАЦИИ ..................................................... |
151 |
| Табель о рангах ................................................................................................................................... |
153 |
| Касты и разделение труда ................................................................................................................. |
167 |
| Жизненный путь ............................................................................................................................... |
187 |
| Государства и федерации .................................................................................................................. |
201 |
| В КРУГУ СЕБЕ ПОДОБНЫХ ........................................................................................................... |
215 |
| Общественный желудок .................................................................................................................... |
217 |
| Внимание к ближнему ....................................................................................................................... |
227 |
| Способы общения ............................................................................................................................. |
236 |
| Визитные карточки ........................................................................................................................... |
249 |
| Мобилизация ...................................................................................................................................... |
258 |
| Хлеб насущный .................................................................................................................................. |
277 |
| МЕЖВИДОВЫЕ ОТНОШЕНИЯ ...................................................................................................... |
291 |
| Место под солнцем ........................................................................................................................... |
293 |
| Мой дом — моя крепость ................................................................................................................. |
306 |
| Воришки, нахлебники и паразиты ................................................................................................... |
320 |
| Узурпаторы и рабовладельцы .......................................................................................................... |
328 |
| ИНТЕЛЛЕКТУАЛЬНЫЕ СПОСОБНОСТИ И ГРУППОВОЕ
ПОВЕДЕНИЕ ...................... |
349 |
| Границы интеллекта ...................................................................................................................... |
351 |
| Феномен коллектива ...................................................................................................................... |
366 |
| На уровне сверхорганизма ............................................................................................................. |
379 |
| Заключение ........................................................................................................................................ | 396 |
| Рекомендуемая литература ................................................................................................................ |
398 |
| Алфавитно-систематический список русских и латинских названий насекомых ....................... | 399 |