| Главная | Библиотека | Форум | Гостевая книга |
В КРУГУ
СЕБЕ ПОДОБНЫХ

Так царя своего ни в Египте не чтут, ни в обширной Вергилий. Георгики |
ОБЩЕСТВЕННЫЙ ЖЕЛУДОК
Как только оса-нянька раздаст принесенный комочек пищи, она приближается, быстро вибрируя крыльями, к отверстию каждой ячейки для того, чтобы выпить слюну, которая обильно струится изо рта заключенной в ней личинки... Но если секрет не появляется, оса захватывает голову личинки челюстями, тянет ее к себе, а затем внезапно вдавливает обратно в ячейку, опуская туда и свою голову. Эти движения стимулируют ротовые органы личинки и заставляют ее выделить слюнную жидкость. Э. Рубо. Биологические исследования одиночных и общественных ос Африки |
Р. Реомюр в шестом томе «Мемуаров...», увидевшем свет в 1742 г., описал удивительное явление, к которому неоднократно обращалась наука в будущем. Наблюдая за обычными во Франции осами-полистами, он заметил, что всякий раз, когда оса накормит личинку, та выделяет изо рта капельку жидкости, которую взрослая оса поглощает как бы в награду. Уже знакомый нам Э. Рубо позднее подробнее исследовал это явление, назвав его экотрофобиозом, что в переводе с греческого прибли-
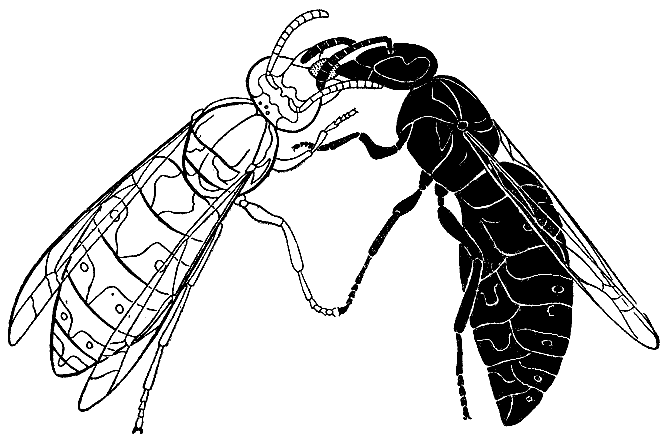
63. Кормовой обмен (трофаллаксис) между двумя рабочими германской осы
(по Г. Монтанье, 1966 г.)
Донор (слева), раздвинув мандибулы, выделяет
капельку корма из зобика. Реципиент
(справа) поглощает эту жидкость, непрерывно постукивая
усиками по голове
и
челюстям донора.
217
|
зительно означает «семейное пищевое сожительство». Он считал, что
это своеобразная форма эксплуатации взрослыми осами личинок
и что именно
эти капельки, получаемые
от личинок, являются основной силой, удерживающей самок в гнезде
и заставляющей их заботиться о потомстве, т. е. в экотрофобиозе
заключена основная причина
возникновения эусоциального поведения. В 1918 г. В. Вилер предложил
для такого явления более удобный и получивший затем широкое
распространение термин трофаллаксис,
что означает «обмен пищей». Он отвергал представления Рубо
об эксплуатации осами личинок и считал, что это, напротив, взаимовыгодная
кооперация
между взрослыми особями и потомством. Действительно, ведь осы
кормят личинок
и дают им гораздо больше пищи, чем получают от них. К началу
XX в. ученым
уже
были известны многочисленные наблюдения взаимного кормления
у
общественных ос, пчел, муравьев, термитов. Поэтому Вилер рассматривал
явление
шире,
называя трофаллаксисом не только кормление личинками взрослых
ос, но и любые формы
передачи жидкой пищи между общественными насекомыми. Короче
говоря, трофаллаксис — это кормление у насекомых изо рта в рот. Партнером
в трофаллаксисе
может быть и личинка. Насекомое, отдающее корм, называют донором,
а поглощающее его — реципиентом.
Следуя за Рубо, Вилер рассматривал трофаллаксис как силу, организующую
насекомых в общество, как основную причину возникновения эусоциальности.
Но, как мы
увидим ниже, трофаллаксиса нет у многих наиболее примитивных
общественных насекомых — стеногастрин, галиктов, аллодапин,
шмелей, ряда низших
муравьев. Значит, эусоциальность вполне может сформироваться
и без трофаллаксиса.
Среди общественных ос трофаллаксис не наблюдали только у стеногастрин,
а все остальные виды, даже такие примитивные
218
полибии, как белоногастер, широко практикуют обмен жидкой пищей
между взрослыми и поглощение личиночных выделений. Во времена
Вилера не
был известен состав
жидкости, получаемой осами от личинок, и поэтому ученые
были вынуждены ограничиваться лишь домыслами и догадками. Исследованиями
в середине
60-х годов установлено,
что личинки ос обладают чрезмерно развитыми по сравнению
с личинками других насекомых нижнегубными или слюнными железами,
которые
и выделяют поглощаемый
осами секрет. Как оказалось, в нем содержится около 9%
сахаров. Выделения, получаемые осами от личинок, нередко составляют
больше половины их
суточного рациона, дополняя нектар, собираемый на цветках,
и другую пищу. Осы частенько
специально выпрашивают корм у личинок, прикасаясь усиками
к их голове. В ответ на такое прикосновение (его можно имитировать
даже кисточкой)
личинка выделяет капельку сладкого секрета. Ранним утром
осы «заправляются» личиночным
секретом и вылетают из гнезда за кормом. Крупная личинка
за одно
«доение» отдает столько сахара, что его хватает для поддержания
жизнедеятельности осы до полудня. Особенно важен этот секрет
для ос в плохую погоду,
когда на цветках провизии не найти, а на обогрев гнезда
энергии нужно много.
Кроме сахаров в личиночном секрете содержится немного ферментов,
расщепляющих белки и углеводы. Их всего около 0,5%, но
и в таком количестве они
действуют достаточно эффективно. Осы смешивают личиночный
секрет с пережевываемой
добычей, и его ферменты подготавливают пищу к усвоению
личинками. Слюнные железы взрослых
ос выделяют очень мало протеаз, и этот недостаток восполняют
ферменты слюны личинок. Получается в некотором роде разделение
функций:
личинки поставляют
ферменты, а осы совершают механическую работу по пережевыванию
добычи и пропитке ее ферментами и дают личинкам в готовом
к проглатыванию виде!
Все эти факты вместе подтверждают точку зрения Вилера,
увидевшего в трофаллаксисе между осами и личинками выгодную
обеим сторонам
кооперацию. Не исключено,
что выделения личинок несут и еще одну функцию: они позволяют
избавляться от излишней влаги в гнезде. Ведь выделение
жидких экскретов обычным
для насекомых путем (через мальпигиевы сосуды и анальное
отверстие) могло
бы привести к
намоканию и порче бумажных сотов.
Примитивные муравьи рода амблиопоне и некоторые другие
понерины никогда не обмениваются кормом. У нотомирмеции,
мирмеций
и большинства понерин
трофаллаксис
наблюдается редко и протекает недостаточно организованно:
позы муравьев при обмене кормом могут быть самыми различными
(т.
е. не стереотипными,
как у
высших муравьев), кормовые контакты непродолжительны. У
высших же муравьев трофаллаксис —
219
обычнейший элемент поведения. Поэтому уже давно сделан вывод, что степень развития трофаллаксиса является одним из важнейших показателей продвинутости социальной организации вида. Но нет правил без исключения, и эта закономерность иногда нарушается в эволюции: у ряда высших муравьев трофаллаксис отсутствует. Примером могут служить муравьи родов афеногастер, мессор, погономирмекс. Муравьи довольно примитивного рода афеногастер, вероятно, никогда не имели трофаллаксиса. А вот у жнецов он не появился, наверное, потому, что пища этих муравьев — в основном твердые зерна и насекомые. Муравьи-кочевники питаются тоже исключительно твердой пищей. Их личинки и царица никогда не получают
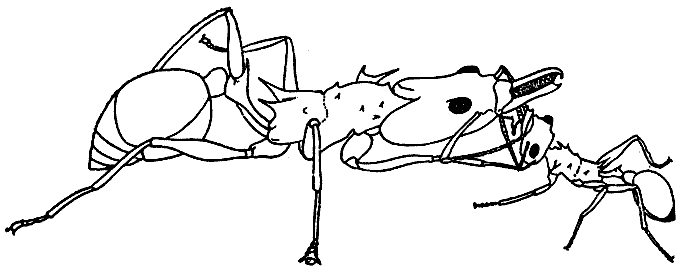
65. Трофаллаксис между рабочими муравья дацетон армигерум (по Э. Вильсону, 1962 г.).
Более крупный рабочий выпрашивает пищу, дотрагиваясь
передними ногами до головы
маленького, и поглощает появившуюся изо рта партнера
жидкость.
корм изо рта рабочих. А между рабочими кормовые обмены
наблюдаются крайне редко. Значит, возможно все-таки
достижение весьма
высокой ступени социальной
организации, на которой находятся кочевники, без
развития трофаллаксиса.
Вообще особенности питания муравьев существенно
влияют на встречаемость трофаллаксических контактов
в их
колониях. Так, почти все формицины
и долиходерины — заядлые
«пастухи»: падь тлей и других сосущих насекомых
занимает в их диете важнейшее место. Некоторые
виды питаются
исключительно жидкой пищей.
Потому-то
у муравьев этих двух подсемейств трофаллаксис —
важнейшая форма
поведения. А вот в пищевом
рационе муравьев подсемейства мирмицин жидкая пища
составляет небольшую долю, даже тлей эти муравьи
посещают не очень
активно,
потому и
трофаллаксис мы
можем наблюдать в их гнездах не столь часто.
Многие ученые видели, как личинки муравьев выпускают
изо рта капельки жидкости, которые слизывают рабочие.
Нередко
муравьи
даже выпрашивают
эти капельки,
постукивая антеннами по ротовым органам и голове
личинки. Царица амблиопон челюстями сдавливает
шею личинки,
заставляя ее выделить
изо рта порцию
секрета, которую затем поглощает. К со-
220
жалению, до сих пор никто не исследовал состав
продуцируемой личинками жидкости. Скорее всего,
это продукт слюнных
желез. Но пока мы ничего
не можем сказать
о его роли в жизни муравьев.
Точно так же недостаточно ясным остается пока
значение капелек, выделяемых личинками из анального
отверстия.
Их тоже поглощают
рабочие. В большинстве
случаев это, конечно, просто экскреты из мальпигиевых
сосудов (иногда хорошо видно, как в капельке
плавают кристаллы
мочевой кислоты).
Но не исключено,
что в них могут быть и какие-то необходимые
муравьям вещества. Не так давно в анальных
выделениях
личинок фараонова муравья
были обнаружены
аминокислоты
и ферменты протеазы. Это позволяет предполагать,
что анальная жидкость личинок может содержать
не только
вредные продукты
обмена веществ.
Среди общественных пчел трофаллаксиса нет у
аллодапин и галиктов. У большинства шмелей
исследователям
не удавалось заметить
обмен кормом, лишь у некоторых
видов крайне редко наблюдали неупорядоченные
трофаллаксисы. Что же касается безжалых и медоносных
пчел, то
трофаллаксис, является
у
них
важнейшим
элементом поведения.
У термитов, как уже говорилось во втором разделе,
две формы трофаллаксиса: оральная, когда пища
выделяется изо рта,
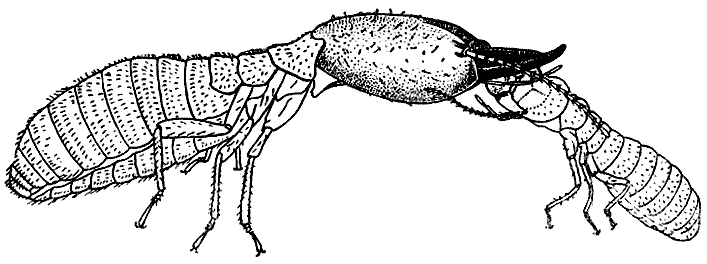
66. Маленький рабочий термита-грибовода макротермес
белликозус кормит
крупного солдата, выделяя
жидкую пищу изо рта
(по П. Грассе, 1949
г.).
и анальная, основанная на выведении корма из заднепроходного отверстия. Анальный трофаллаксис есть только у низших термитов и служит в первую очередь для передачи кишечных симбионтов. Кроме того, этим путем полупереваренная пища из толстой кишки одного термита окончательно усваивается другими членами колонии. При оральном трофаллаксисе рабочие термиты могут выделять корм двух типов. Секрет слюнных желез рабочие дают царской паре, крылатым термитам и личинкам самых младших возрастов; он составляет единственную пищу этих насекомых. Более старшим личинкам, нимфам, солдатам и другим рабочим они отрыгивают содержимое зобика или средней кишки, т. е. частично переваренную пищу. У выс-
221
ших термитов секретом слюнных желез рабочих
питаются личинки всех возрастов.
При трофаллаксисе насекомые принимают
стереотипные позы, характерные для
семейства, рода и
даже вида. Мы еще поговорим
об этом,
когда речь пойдет
о способах общения. Если существует
поведенческая иерархия, то корм всегда
передается от
подчиненного насекомого
доминирующему; донора
и реципиента
легко различить. У пчел при трофаллаксисе
донор лишь слегка выдвигает
хоботок вперед и вниз и выделяет из
него каплю корма, а пчела-реципиент
почти
полностью вытягивает хоботок вперед,
высасывая эту
каплю.
А вот у муравьев или термитов различить
донора и реципиента
не
всегда просто,
если только
не было возможности
увидеть, кто именно из партнеров отрыгнул
каплю корма. Впрочем, у целого ряда
видов муравьев
решить эту
задачу
несложно,
поскольку донор широко
раскрывает челюсти, а реципиент держит
их сомкнутыми.
Недавно во Франции была разработана
установка, позволяющая измерять радиоактивность
отдельных частей тела крупных
муравьев в процессе
трофаллаксиса. Когда
рабочих кампонотуса накормили раствором
сахара с радиоактивным золотом, то
удалось
при обменах кормом между ними по уменьшению
радиоактивности брюшка донора и увеличению
ее у реципиента измерять
скорость трофаллаксиса.
На этой
же установке было обнаружено и ранее
неизвестное явление — реверсии трофаллаксиса.
Выяснилось,
что нередко в
процессе обмена кормом
донор и реципиент вдруг
меняются ролями и корм начинает поступать
в зобик того муравья, который только
что его
отдавал.
Иногда в процессе
одного
трофаллаксиса
роли
муравьев могут
поменяться даже несколько раз. Пока
непонятно, какой смысл заключен в подобных
реверсиях.
Основная функция трофаллаксиса — распределение
пищи между всеми насекомыми в колонии.
Важнейшая особенность
системы
такого
распределения — ее
открытость: каждый рабочий, солдат,
личинка, крылатая самка или самец могут
вступить
в обмен кормом с любым членом колонии.
Корм обычно отдают по первому требованию
и получают
при необходимости
от
других. Поэтому совокупность
зобиков всех
насекомых в гнезде составляет как бы
общественный желудок, в котором корм
хранится и перераспределяется
путем
трофаллаксиса. Впервые
представление об общественном желудке
ввел в науку О. Форель в 1878 г.
У видов, не обладающих трофаллаксисом,
нет и общественного желудка. Некоторое
исключение
в
этом отношении
составляют только шмели.
В их гнезде есть
медовый горшок — временное хранилище
резервных запасов меда. Шмели то пополняют
запасы, то используют их. Когда одному
шмелю скормили немного радиоактивного
сиропа,
следы радиоактивности вскоре
222
обнаружились у многих особей из этого гнезда, поскольку они, как говорится, «хлебали из одной лохани» — медового горшка. Не все виды общественных насекомых в равной мере способны сохранять в общественном желудке резервные запасы корма. В значительной степени это обусловлено строением пищеварительной системы. У всех насекомых зобик представляет собой расширение передней кишки и связан с ротовым отверстием пищеводом. Следующим отделом кишечника является мышечный желудок, или провентрикулюс. У термитов он снабжен зубчиками, которые дополнительно перетирают проглатываемый
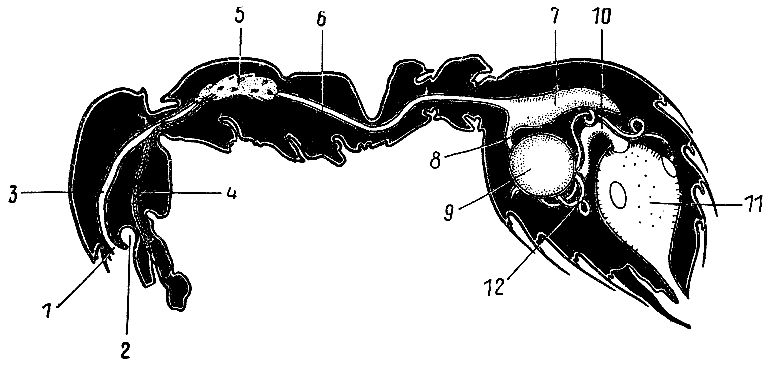
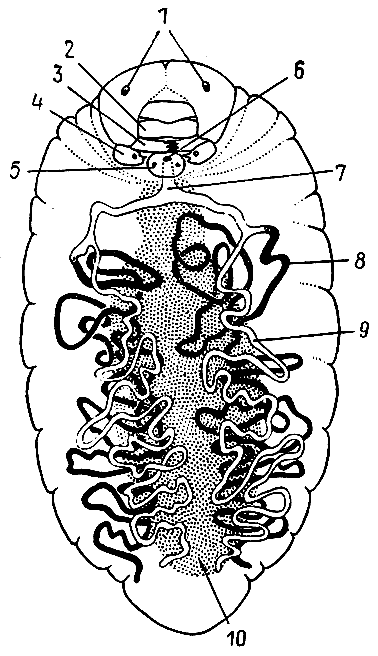
67. Пищеварительная система рыжей мирмики (по Ш. Жанэ, 1894 г.).
1 — ротовая
полость; 2 —
подротовой карман;
3 — глотка;
4 — проток
слюнной
железы;
5 — слюнная железа; 6 — пищевод;
7 — зобик; 8 —
мышечный желудок (провентрикулюс);
9 — железистый желудок;
10 — толстая кишка;
11 — прямая кишка; 12 —
выделительные
трубочки (мальпигиевы сосуды).
корм. У перепончатокрылых единственная
функция провентрикулюса — управление
поступлением пищи из зобика
в собственно желудок. Зобик
покрыт изнутри
кутикулой, поэтому пища в нем
не переваривается и не усваивается.
Значит, когда пчела
или муравей проглатывает корм,
то он попадает
как бы в общественный желудок
и
пока не является
«собственностью»
данного насекомого.
Ведь из зобика
корм по первому требованию
может быть отдан любому другому. Выходит,
что,
так сказать, «индивидуальным
ртом» муравья,
термита или пчелы служит провентрикулюс,
ведь насекомое
питается,
только
когда клапаны
мышечного желудка открываются
и пища поступает в собственно
желудок.
У всех низших муравьев, а также
у мирмицин провентрикулюс устроен
так,
что просвет
его закрывается сокращением
кольцевой мышцы.
Для того чтобы
держать
пищу в зобике, муравей должен
непрерывно напрягать ее. Видимо,
по этой причине
названные муравьи
не могут длительное
время
хранить в общественном
желудке
значительные запасы пищи.
223
У формицин и долиходерин провентрикулюс
имеет совершенно иное строение:
его хитиновые клапаны
постоянно
плотно сомкнуты и
открываются только
при сокращении
специальных мышц. Поэтому
хранение пищи в зобике не требует особых
усилий. Виды
муравьев
этих
двух подсемейств
обладают
мягким растяжимым
брюшком
и способны приносить в
гнездо и удерживать в зобиках намного
больше
пищи, чем
все прочие муравьи. Длительное
хранение в гнезде жидкой
пищи у них весьма
обычное явление.
Стоит
дать содержащимся
в лаборатории
формикам
или лазиусам
раствор сахара либо меда,
как среди них окажется множество
рабочих
с сильно раздутыми
брюшками.
Зобики их до отказа
заполнены сладким
кормом.
Эти распухшие
муравьи могут в течение
многих недель хранить внутри себя
общие запасы
корма. Именно
среди формицин
и долиходерин встречаются
так называемые
медовые муравьи,
в гнездах которых можно
найти «медовые бочки» — рабочих
с полупрозрачным брюшком, раздувшимся
от сладкой
жидкости до размера крупной
вишни. Их называют
плерэргатами.
Американские медовые муравьи
принадлежат к роду мирмекоцистус
и обитают
в прериях. В их
глубоких
почвенных гнездах
можно найти до 1500 плерэргатов.
Каждый сохраняет около
0,4 г «меда», что составляет
до 90%
массы его
тела. Запасы
меда в муравейнике
могут достигать
600 г. Муравьи
выходят
из гнезд
по ночам и собирают падь
тлей, а затем отдают ее
плерэргатам,
которые
неподвижно висят, уцепившись
ножками или челюстями за
стенки
глубоких
камер
муравейника.
При этом зобик их так раздут,
что спинные и
брюшные полукольца занимают
на брюшке совсем небольшую
площадь,
остальное — это межсегментарная
мембрана. Когда
возникает потребность,
плерэргаты отрыгивают
корм любой попросившей
его особи. Так и проходит
вся жизнь муравья-бочки,
она не
балует его разнообразием!
Муравьиный мед не очень
сладкий (содержит не более
50% сахаров),
но, как и
пчелиный, включает
в основном
простые
сахара и
оксидазу глюкозы,
стерилизующую
мед за счет выделения перекиси
водорода. Некоторые плерэргаты
хранят почти
чистую, слегка подслащенную
воду. Запас
воды в тех засушливых
районах, где живут медовые
муравьи, тоже очень кстати!
В сухих районах Австралии
обитают муравьи кампонотус
инфлатус,
плерэргаты которых
почти столь же
огромны, как у американских
медовых муравьев,
а также виды родов мелофорус
и лептомирмекс, обладающие
«медовыми бочками» с менее
растяжимым брюшком. Муравьи
с такими
плерэргатами встречаются
и в других районах мира:
это
тапинома в Южной Америке,
аноплолепис и
плагиолепис
в
Африке, проформика и пренолепис
в Азии. На юге СССР живет
несколько видов рода
проформика.
В гнездах этих муравьев
постоянно есть плерэр-
224
гаты. Брюшко их увеличено
не очень сильно, так
что плерэргаты
свободно
ходят по
гнезду. Но все
же в
зобике каждого
хранится около 10 мг
сладкого корма,
а этого достаточно
для питания сотни муравьев
в течение
месяца.
Скорость перемешивания
пищи в общественном
желудке удивительно
велика. Когда
шести пчелам скормили
20 мл сиропа с радиоактивным
фосфором,
то через пять
часов 27% рабочих в
улье
имели радиоактивную
метку, а через
сутки их было уже
55%. Значит, всего
за сутки принесенный
шестью фуражирами
корм попал
в зобики десятков тысяч
пчел. Сколько же потребовалось
для этого актов
трофаллаксиса! В подобном
же эксперименте один
рабочий бурого
лесного муравья
набрал полный
зобик сиропа с радиоактивным
йодом. Через 27 часов
все 70 муравьев,
обитавших в
лабораторном гнезде,
были радиоактивными.
Многочисленные
опыты с весьма
сходными результатами
неоднократно проводились
и с другими
видами
общественных пчел,
ос, муравьев,
термитов.
Но в то же время эксперименты
обнаружили и некоторые
различия. Виды, в колониях
которых обмен кормом
происходит в среднем
реже, отличаются,
как
и следовало
ожидать, меньшей скоростью
перемешивания пищи
в общественном желудке.
Например,
у мирмицин принесенный
в гнездо
корм значительно медленнее
распределяется
между рабочими, чем
у формицин или долиходерин.
Различия
в скоростях перемешивания
корма
могут быть связаны
и с его природой.
Доставленная
фуражирами рыжих
лесных муравьев падь
очень быстро
распределяется в основном
среди внегнездовых
рабочих,
а гемолимфу добычи
фуражиры
передают прежде
всего рабочим-нянькам
для личинок, и она
распространяется
среди муравьев в гнезде
в 2—3 раза медленнее,
чем сладкий
корм.
Не все насекомые в
колонии совершенно
равноценны
в системе трофаллаксиса.
Царицы обычно
только получают корм,
но
никому не отдают его.
Впрочем, у примитивных
видов общественных
насекомых царицы
могут иногда
выступать в роли доноров.
Такое наблюдается даже
у некоторых муравьев,
в частности
у мирмик.
Крылатые самки
и самцы, солдаты,
личинки
я нимфы также
чаще являются
реципиентами.
Значение трофаллаксиса
не ограничивается распределением
пищи и перемешиванием
ее в общественном желудке,
хотя это, несомненно,
наиболее важная
его функция. Огромна
его роль и в регуляции
потребления
пищи
и
фуражировочной деятельности
насекомых. Спрос в
колонии на пищу каждый
ее член
ощущает,
так сказать,
на «собственном
брюхе». Когда
корма не
хватает, зобик
фуражира пустеет,
и это
прибавляет ему энергии
в поисках добычи.
Трофаллаксис может
применяться для передачи
информации.
Например, при
большой потребности
в пищи фуражира
«атакует» сразу несколько
рабочих,
выпрашивающих
у него
225
корм. Это служит сигналом
к усилению фуражировочной
деятельности.
При мобилизации
на пищу пчелы и муравьи
нередко
используют
трофаллаксис для
передачи информации о качестве
корма.
Еще одна функция
трофаллаксиса связана
с распространением
в колонии феромонов.
Установлено,
что феромоны
царской пары попадают
ко
всем термитам именно
при трофаллаксисе,
вместе с пищей.
Существует предположение,
что
таким же путем
могут переходить
от
особи к
особи феромоны
и
у общественных
перепончатокрылых.
Во всяком случае,
трофаллаксис
значительно ускоряет,
например, передачу
феромона
пчелиной
матки, поскольку
способствует более
тесным контактам
между
рабочими,
во время которых
феромон как
бы «перетекает»
с одной пчелы на
другую.
Голодные личинки
ос, выделяя капельки
сладкой
слюны,
привлекают этим
рабочих ос. Расчет
здесь простой:
оса выпьет капельку,
обратит внимание
на голодную
личинку да и покормит
ее! В этом случае
трофаллаксис обеспечивает
функцию привлечения.
И, наконец, трофаллаксис
может снижать агрессивность
при враждебных
столкновениях.
Вспомним, как
подчиненная оса
выделяет капельку
корма и этим «усмиряет»
доминирующую. Так
бывает и у муравьев,
но уже
в территориальных
взаимоотношениях.
Когда
муравей случайно
или целенаправленно
попадает во владения
соседней колонии
и
вдруг оказывается
окруженным
несколькими
враждебно
настроенными чужаками,
он поступает весьма
находчиво: отрыгивает
капельку
корма и предлагает
ее врагам. И чем
агрессивнее они
себя ведут,
тем
настойчивее
прельщает их
кормом атакованный
муравей.
Нередко это спасает
храбреца. Враги
поглощают пищу
и
явно «добреют»,
начинают чиститься,
не обращая внимания
на
нарушителя
границы, который
успевает
улизнуть.
Такое поведение
наблюдали
даже между муравьями
разных видов. Настоящий
межвидовой трофаллаксис!
Как могло возникнуть
в эволюции столь
сложное поведение
—
трофаллаксис? Вероятнее
всего, это произошло
на основе случайного
вначале
слизывания капелек
пищи, попавших
на тело фуражиров.
Потом насекомые
могли
уже
облизывать
более
целенаправленно
не только
тело, но
и ротовые органы,
на которых были
остатки корма.
Отсюда недалеко
и до специального
отрыгивания
корма донором и
поглощения
его реципиентом.
Сначала такой кормовой
обмен
был неупорядоченным,
как
у некоторых современных
примитивных муравьев.
Затем постепенно
сформировались
стереотипные позы
выпрашивания, передачи
и приема корма,
а это значительно
повысило результативность
всего
процесса. Косвенным
подтверждением
такого
эволюционного
пути является тот
факт, что трофаллаксис
есть
только в тех группах
общественных
насекомых, где
практику-
226
ется взаимное облизывание. Исключение составляют осы-полисты, у которых взаимное облизывание наблюдается очень редко, но трофаллаксис весьма обычен.
ВНИМАНИЕ К БЛИЖНЕМУ
Как у коров, взаимно облизывающих друг друга, так и у муравьев облизывание сотоварищей представляет собой деятельность, большей частью приятную для облизываемой части тела. Э. Васманн. Итоги сравнительной психологии |
Очистка
и облизывание
собственного
тела —
важнейшая
форма
поведения
насекомых.
Она обеспечивает
удаление
болезнетворных
микроорганизмов,
остатков
пищи и
прочих
частиц,
загрязняющих
поверхность
тела, мешающих
движениям
и работе
органов
чувств.
Все насекомые
очень часто
чистятся.
Но у наиболее
высокоорганизованных
общественных
насекомых
к
«индивидуальному
туалету»
добавилась
взаимопомощь
в поддержании
чистоты
тела —
груминг (от англ.
to groom
—
«чистить
лошадь»),
или аллогруминг (от греч.
allos —
«иной,
другой»),
т.е. облизывание
и очистка
тела другой
особи.
У примитивных
эусоциальных
насекомых
аллогруминга
нет. Рабочие
шмели,
галикты,
осы обычно
не проявляют
специального
внимания
друг к
другу.
Взаимодействия
между ними,
если не
считать
контактов
доминирования-подчинения,
непродолжительны
и ограничиваются
ощупыванием
усиками
или
случайными
прикосновениями;
осы, помимо
этого,
вступают
еще в трофаллаксис.
Аллогруминг
у
ос-полистов
можно наблюдать
крайне
редко,
обычно
при попадании
на тело
капелек
сладкой
пищи
или иных
веществ.
Даже
у таких
достаточно
высокоорганизованных
в социальном
отношении
ос, как
веспины,
это поведение
— редкость.
И в гнездах
безжалых
пчел нечасто
удается
быть свидетелем
аллогруминга.
А вот
у муравьев,
медоносных
пчел и
термитов
это один
из характернейших
элементов
поведения
внутри
гнезда.
Если понаблюдать
за муравьями
или термитами
в лабораторном
гнезде
и за медоносными
пчелами
в стеклянном
улье, то
можно убедиться,
что аллогруминг
встречается
у них значительно
чаще, чем,
например,
трофаллаксис.
Это
регулярное,
каждодневное,
так сказать,
рутинное
занятие
сотен
и
тысяч насекомых.
Очень забавно
видеть,
как рабочий
тщательно
и долго
вычищает
и вылизывает
язычком
тело
своего
собрата,
методично
переходя
от головы
к
груди,
затем к
ножкам,
брюшку
и т.д.,
а
насекомое,
которое
чистят,
приподнимает
то одну
нож-
227
ку, то
другую,
поворачивает
голову,
брюшко,
меняет
позы,
подставляя все новые
участки
тела.
Если бы по отношению
к насекомым
были
допустимы подобные
выражения,
то
следовало бы сказать,
что
чистильщик исполнен
сознанием
важности
и ответственности
за
выполняемое дело,
а подвергаемое
чистке
насекомое
явно
не
скрывает получаемого
удовольствия
и
стремится
его
продлить!
Но
если
оставить
эмоции,
то
нужно
подчеркнуть,
что
аллогруминг
имеет
важнейшую
социальную
функцию
—
поддержание
чистоты
тела
и работоспособности
всех
членов
колонии.
Поэтому
он
особенно
проявляется,
когда
«правила
гигиены»
нарушаются.
Например,
фуражиры
принесли
в гнездо
много
жидкой
пищи
и при
интенсивных
трофаллаксисах
тела
многих
насекомых
загрязнились
кормом.
Вот
тут-то
и
происходит
массовое
наведение
чистоты:
рабочие
чистятся
сами
и
тщательно
вылизывают
друг
друга.
Нередко
можно
видеть,
как
одного
рабочего
чистит
сразу
несколько
его
собратьев.
Многие
ученые
предполагают,
что
аллогруминг
способствует
распространению
в колонии
феромонов,
находящихся
на
поверхности
тел
насекомых,
и что
именно
в этом
и состоит
его
основная
социальная
функция.
Однако
экспериментальных
доказательств
такого
предположения
пока
нет.
У примитивных
муравьев
— мирмеций,
понерин
— аллогруминг
можно
наблюдать
значительно
реже,
чем
у представителей
более
высокоорганизованных
подсемейств.
А вот
среди
термитов
—
как
низших,
так
и
высших
— он
встречается
в равной
степени
часто.
У этих
насекомых
аллогруминг
играет
еще
одну
важную
социальную
роль.
Дело
в
том,
что
при
линьках,
т.
е.
при
сбрасывании
старой
кутикулы,
все
термиты:
личинки,
нимфы,
рабочие
— нуждаются
в
посторонней
помощи.
Самостоятельно
перелинять
термит
обычно
не
может
и погибает,
так
и не
выбравшись
из
отслужившей
свое
шкурки.
Вот
тут-то
и приходят
на
выручку
рабочие
термиты,
постоянно
занимающиеся
этим
важнейшим
делом.
К.
С.
Шатов
назвал
их
грумами.
Термиты,
собирающиеся
линять,
ложатся
в глубине
гнезда
на
бок
и,
сильно
выгибая
спину,
сворачиваются
«калачиком».
Вскоре
шкурка
их
лопается
на
спинке
и начинается
линька.
В
этот
момент
к термиту
обязательно
подходят
грумы
и,
тщательно
вылизывая
его,
помогают
освобождаться
от
шкурки.
Когда
все
закончится
благополучно,
грумы
вылижут
все
тело
перелинявшего
термита,
и он
может
отправляться
по
своим
делам.
У примитивных
эусоциальных
насекомых
царица
обычно
«внушает
страх»
рабочим
и они
не
проявляют
к ней
«нежных
чувств»,
только
кормят,
да
и то
после
некоторого
принуждения
с ее
стороны
в
форме
доминирования
или
прямой
враждебности.
Но
у
эволюционно
продвинутых
общественных
насекомых,
когда
доминирование
уже
выражено
незначительно
228
или
совсем исчезло,
царица становится
привлекательной для
рабочих и
возникают различные
формы ухода
за ней.
У царицы теперь
есть свита
рабочих. Так,
постоянно окружающие
царицу восточного
шершня рабочие
слизывают с
поверхности ее
тела выделяемый
из мандибулярных
желез феромон.
Осы делают
это столь
тщательно, что
постепенно сдирают
язычками все
опушение, и
в результате бока
головы, груди
и брюшка
старой царицы
становятся блестящими.
Рабочие свиты
кормят царицу
и повсюду следуют
за ней.
У цариц ос
рода веспула
свиты никогда
не бывает.
Царицу
медоносных пчел
обычно окружает
свита из
5—10 молодых
рабочих с
развитыми кормовыми
железами. Пчелы
стоят головами
к царице,
образуя вокруг
нее кольцо.
Время от
времени они
прикасаются к
царице усиками,
а иногда
облизывают ее
тело. Каждые
10—20 минут
пчелы кормят
царицу, отрыгивая
ей в
рот молочко.
Когда царица
движется по
сотам, свита
следует за
ней и
вновь окружает
кольцом, как
только она
остановится. Свита
не имеет
постоянного состава
(это, видимо,
характерно для
всех общественных
насекомых), и
каждая пчела,
поработав в
ней некоторое
время, уходит
и начинает
заниматься другими
делами, ее
сменяют новые
пчелы.
Царицы
безжалых пчел
в разной
степени привлекательны
для рабочих.
У пчел
одних видов
(мелипоны) превалирует
страх перед
царицей и
свиты практически
нет. У
других же
(паратригоны) множество
рабочих окружает
царицу, дает
корм, облизывает
ее тело
и особенно
кончик брюшка.
У большинства
видов безжалых
пчел царица
питается только
из заполняемых
провизией ячеек
и кормовыми
яйцами рабочих,
но у
некоторых получает
пищу и
при трофаллаксисе
с рабочими.
Интересно, что
пчелы никогда
не предлагают
царице корм
сами, а
лишь отвечают,
да и
то далеко
не всегда,
на выпрашивание
с ее
стороны.
В
эволюции многих
групп общественных
насекомых заметна
немаловажная тенденция
— возрастание
привлекательности царицы
для рабочих,
зависимости ее
от них
и уменьшение
самостоятельности. Проследим
это
на примере
муравьев.
Царицы
мирмеций и
большинства понерин
по поведению
мало отличаются
от рабочих
муравьев и
даже нередко
выходят из
гнезда за
кормом. Свиты
у них
не бывает.
У мирмик
царицы тоже
весьма независимы:
они бродят
по всему
гнезду, питаются
принесенной рабочими
добычей и
даже ухаживают
порой за
личинками. Совершенно
самостоятельно откладывает
царица и
яйца: подгибая
брюшко вперед,
она принимает
появляющееся яйцо
челюстями и
помещает его
в пакет.
Лишь иногда
яйца относят
в пакет
рабочие. Царицы
мирмик в
небольшой степени
привлекательны для
рабочих и
свиты не
имеют. Муравьи
очень редко
облизывают своих
цариц и
никогда не
предлагают корма.
Приходится царице
самой выпрашивать
пищу у
рабочих, а
зачастую, подобно
самцам общественных
ос, при-
229
страиваться
сбоку от
обменивающихся кормом
рабочих, чтобы
поживиться от
их капельки.
Царица
рыжих лесных
муравьев тоже
достаточно самостоятельна,
но получает
пищу только
от рабочих
и за
личинками никогда
не ухаживает.
Рядом с
ней всегда
бывает несколько
муравьев. Они
облизывают царицу
и предлагают
ей корм.
Но и
сама она
нередко выпрашивает
пищу у
рабочих. Откладывая
яйца, царица
подгибает брюшко
и иногда
берет их,
но чаще
яйцо уносят
рабочие. Подобные
же взаимоотношения
между царицей
и рабочими
можно наблюдать
у кампонотусов,
муравьев-портных и
многих других
видов, у
которых физогастрия
цариц выражена
незначительно и
свита невелика.
При
увеличении физогастрии
царица все
более теряет
самостоятельность. Например,
царица
муравья-вора при
откладке яйца
подгибает брюшко,
но кончик
его не
достигает головы,
и царица
не может
сама взять
яйцо. Его
всегда уносят
рабочие. Сильно
физогастричные царицы
из крупных
семей черного
садового или
желтого земляного
муравьев малоподвижны
и практически
ничем не
занимаются, кроме
питания и
откладки яиц.
Такая царица
неизменно окружена
массой муравьев.
Они толпятся
со всех
сторон и
сидят сверху
на ее
брюшке, груди
и даже
голове. Рабочие
муравьи постоянно
облизывают царицу
и уносят
откладываемые яйца.
Царице не
приходится даже
слегка подгибать
брюшко, да
она и
не смогла
бы этого
сделать. Муравьи
сами предлагают
ей корм,
царица никогда
его не
выпрашивает.
Максимального
развития свита
царицы достигает
у муравьев-кочевников.
Их передвигающуюся
по тропе
физогастричную царицу
окружают сотни
муравьев, среди
которых в
несколько раз
больше крупных
рабочих и
солдат, чем
в остальных
частях колонны.
Ведь основное
назначение свиты
у кочевников
— охрана
единственной царицы.
Муравьи иногда
даже прикрывают
ее сверху
своими телами,
а если
возникает опасность,
образуют вокруг
нее плотный
клубок из
тысяч особей.
Свита вытянута
вдоль тропы,
и ее
авангард опережает
царицу почти
на метр,
а арьергард
теснится на
1—2 м
позади. На
этом протяжении
примерно в
пять раз
больше муравьев
на единицу
площади, чем
в остальной
колонне. Группа
муравьев свиты
не является
постоянной. Если
царица останавливается,
колонна продолжает
движение и
свиту образуют
муравьи, которые
до этого
шли сзади.
Очень
сходную картину
мы наблюдаем
у термитов.
Царь и
царица низших
термитов достаточно
самостоятельны, свободно
перемещаются из
камеры в
камеру, оставаясь,
впрочем, в
глубине гнезда.
Они привлекательны
для термитов,
но свита
их обычно
невелика. Окружающие
царскую пару
рабочие облизывают
царя и
царицу, кормят
их выделениями
слюнных желез
и уносят
откладываемые яйца.
Свита царской
пары большого
230
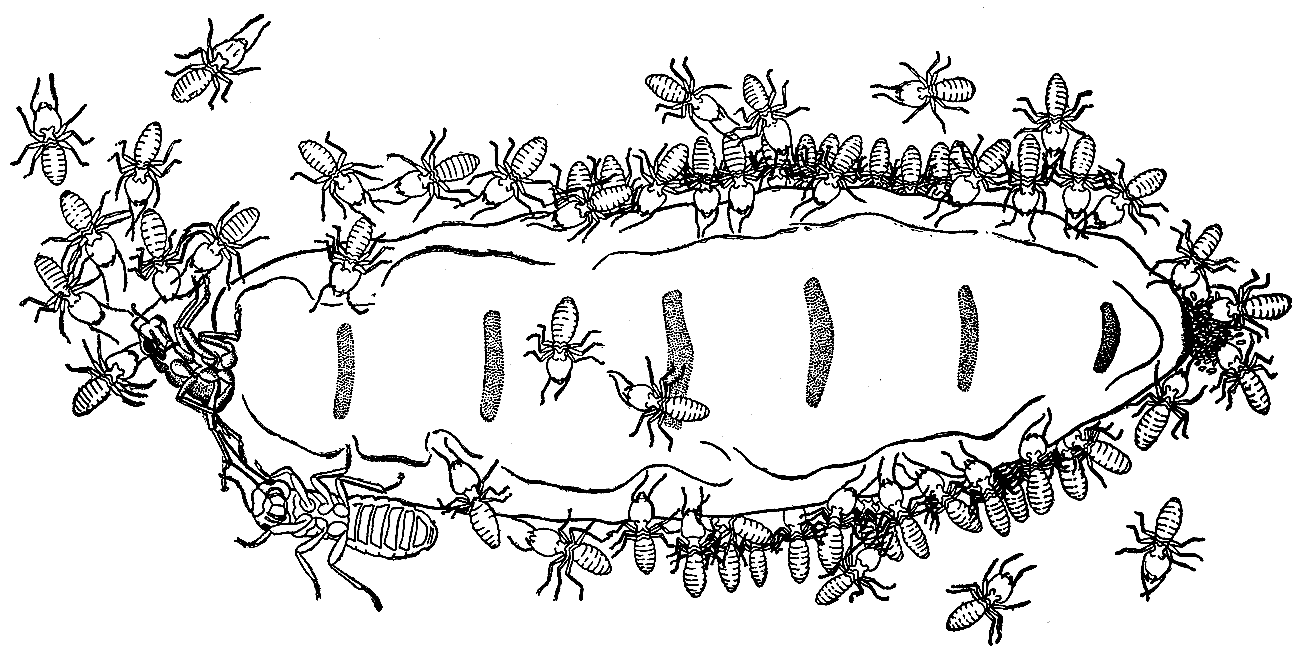
68. Царица термита макротермес белликозус, окруженная свитой рабочих (рисунок сделан снизу через стекло) (по П. Грассе, 1949 г.).
Рабочих особенно привлекает брюшко царицы, которое они старательно облизывают, причем отдельные особи даже забираются под него. У заднего конца брюшка царицы — множество отложенных ею яиц. Неподалеку от головы царицы, возле ее брюшка, можно заметить царя.
закаспийского
термита отчетливо
выражена только
в период
откладки яиц
и состоит
из 10—30
рабочих второго-третьего
возрастов и
одного-двух солдат.
Старшим рабочим,
а также
нимфам и
крылатым почему-то
«запрещено» приближаться
к «монархам»:
оказавшись в
3—4 см
от царской
пары и,
вероятно, ощутив
ее запах,
они резко
разворачиваются
и
убегают.
Физогастрия
цариц высших
термитов,
как
уже говорилось,
может достигать
крайней степени.
Такая царица
совершенно
беспомощна
и всю
жизнь, не
двигаясь,
проводит
внутри царской
ячейки. Сюда
днем и
ночью идут
вереницы рабочих.
Одни толпятся
вокруг головы
грузной мамаши,
наперебой
предлагая
ей пищу.
Другие тщательно
облизывают
ее
огромное брюшко.
Третьи непрерывным
потоком уносят
откладываемые
ею
яйца. Здесь
всегда есть
и солдаты,
но они
«стоят на
страже» неподалеку
и не
изливают своих
«верноподданнических
чувств»
так
рьяно, как
рабочие, стремящиеся
из всех
сил пробраться
к брюшку
царицы и
слизать с
него нечто,
неодолимо
их
притягивающее.
Это
нечто, конечно,
является феромонами
царицы. Рабочие
старательно
вылизывают
поверхность
брюшка,
теребят, даже
слегка прикусывают
складки его
кожи. Нередко
они забираются
на тело
царицы и
покрывают
все
ее брюшко
сверху. Если
царицу со
свитой вынуть
из царской
ячейки и
поместить
в
садок с
почвой, то
термиты первым
делом быстро
построят вокруг
царицы стенки,
затем возведут
крышу —
и укрытие
готово! Царь
тоже имеет
свиту, но
внимания ему
оказывается
значительно
меньше.
Заботой
в гнездах
общественных
насекомых
окружены не
только плодовитые
самки и
самцы, не
только взрослые
солдаты и
рабочие, но
и яйца,
личинки, куколки.
Впрочем, у
большинства
пчел
мы не
увидим сложных
форм ухода
за потомством.
Яйцо лежит
в ячейке
и не
привлекает
особенно
рабочих. Личинка
пчел получает
корм не
в рот,
а просто
на дно
ячейки. Галикты
и безжалые
пчелы вообще
не вступают
в контакт
со своим
потомством,
развивающимся
в запечатанных
ячейках. Исключение
в этом
отношении
представляют
аллодапиновые
пчелы,
но и
их поведение
не сложнее,
чем у
большинства
ос.
Забота о
личинках у
ос ограничена
кормлением.
Правда,
жидкую пищу
осы отрыгивают
личинкам прямо
в рот,
а твердые
«котлетки»
помещают
на ротовые
органы. Но
вся остальная
жизнь личинки
проходит без
участия рабочих:
она растет,
линяет и
окукливается
самостоятельно.
Взрослое
насекомое
также
может выйти
из куколки
без помощи
рабочих.
Иное
дело —
муравьи и
термиты. Даже
яйца этих
насекомых не
могут нормально
развиваться без
ухода —
рабочие постоянно
перекладывают
их,
облизывают и
перетаскивают
с
места на
место. Слюна,
попадающая на
поверхность яиц
при облизывании,
обладает бактерицидными
свойствами. Поэтому
яйца, лишенные
ухода, нередко
покрываются плесенью
и погибают.
232
У
термитов облизывание
яиц выполняет
еще одну
функцию: яйца
впитывают
из
слюны необходимую
для развития
воду и
постепенно разбухают
в объеме
на 30—50%.
Появившиеся
из
яиц личинки
термитов в
передвижениях
вполне
самостоятельны,
но
получают корм
только от
рабочих и
не могут
перелинять без
помощи грумов.
Уход
за потомством
у муравьев
довольно
разнообразен.
Личинки наиболее
примитивных
муравьев
— нотомирмеции,
мирмеций,
амблиопон
и некоторых
других
понерин
— весьма
самостоятельны.
Они
даже способны
к ограниченным
передвижениям.
Например,
эти существа
проползают
1—2
см, чтобы
добраться
до
принесенной
рабочими
добычи, вгрызаются
в нее
и выедают
все мягкие
части.
Когда
личинки голодны,
они порой
даже нападают
на своих
собратьев!
Мелкие
личинки
получают
от рабочих
кусочки насекомых
и кормовые
яйца. Иногда
муравьи
отрыгивают
в рот
личинкам жидкую
пищу, но
такой способ
кормления
играет
у них
небольшую
роль,
а у
амблиопон
вообще
отсутствует,
и
их личинки
вырастают
исключительно
на твердой
пище.
Личинки
всех
остальных
муравьев
совершенно
не могут
передвигаться
самостоятельно
и
тем
более
находить
корм.
Поедают
они
только
специально
обработанную
добычу.
Доставленных
в
гнездо
насекомых
муравьи
раздирают
на части,
поглощая
гемолимфу
жертвы.
При
пережевывании
кусочков
рабочие
выделяют
ферменты
слюны,
которые
начинают
процесс
переваривания,
а образующиеся
соки
всасывают.
В конце
концов
от
жертвы
остаются
только
хитиновые
покровы,
выбрасываемые
из гнезда.
Личинки
младших
возрастов
получают
прямо
в рот
из зобиков
рабочих
гемолимфу
и частично
переваренные
соки
добычи,
а
также
падь
тлей
и
другие
содержащие
сахар
жидкости.
Кроме
того,
высшие
муравьи
дают
личинкам
секреты
кормовых
желез
—
лабиальных
(нижнегубных)
и постфарингеальных
(заглоточных).
Эти
же секреты
получают
и
царицы
муравьев.
В постфарингеальные
железы
муравья
при
пережевывании
кусочков
добычи
попадает
значительное
количество
содержащихся
в них
жиров.
Эти
жиры
рабочие
и отдают
личинкам.
Старшие
личинки
наряду
с жидкой
пищей
получают
пропитанные
ферментами
кусочки
насекомых.
Муравьи
кладут
комочек
на
нижнюю
сторону
тела
личинки
возле
ее
ротовых
органов,
и та
постепенно
поедает
его.
Для
того
чтобы
удерживать
корм,
личинки
многих
видов
имеют
специальные
приспособления.
У личинок
огненного
муравья
это
длинные
прочные
волоски,
окаймляющие
лишенную
опушения
округлую
область
на
нижней
стороне
груди
и
направленные
вперед;
они
поддерживают
кусочек
пищи
сзади
и
с боков.
Личинки
тетрамориумов
имеют
два
поперечных
валика
на нижней
стороне
шестого
и седьмого
сегментов
тела.
Эти
валики
не позволяют
комочку
пищи
ото-
233
|
двинуться
назад,
когда,
пригнув
голову,
личинка
грызет
его
челюстями.
У личинок
лептотораксов
подобные
валики
развиты
на третьем,
четвертом
и
пятом сегментах,
причем
самый
крупный
—
последний.
А
у личинок
кампонотусов
на
том
же
месте расположена
ямка.
Личинка
подгибает
голову
к груди
и постепенно
поедает
кусочек
пищи, который
муравьи
кладут
в эту
ямку.
На
груди личинок
псевдомирмексов
ямка
столь
велика,
что похожа
на карман,
открывающийся
к
голове;
в
него
рабочие
помещают
кусочки
питательных
телец
Белта,
собираемых
на листьях
акации,
и
другую
пищу.
Подобные
приспособления
для удерживания
пищи
очень
разнообразны,
но развиты
только
у личинок
старших
возрастов.
Муравьи
родов
афеногастер,
катаглифис,
лазиус
и
др.
приносят
наиболее
крупных
личинок
к кускам
добычи
и
кладут
на
них.
Личинки
самостоятельно
выедают
все
мягкие
ткани,
напоминая
этим
потомство
примитивных
муравьев.
Два-три
передних
сегмента
тела
у личинок
сужены
в
виде
«шеи»,
что
придает
голове
большую
подвижность
и
позволяет
насекомому
легко
управляться
с кормом.
Иногда
на
куске
добычи
можно
видеть
более
десятка
личинок,
причем
головы
их
всегда
находятся
на
пище,
а брюшки
направлены
в
разные
стороны,
как
лепестки ромашки.
Большинство
муравьев
дает
личинкам
и
кормовые
яйца. Нередко
личинки
первого
возраста
питаются
исключительно
ими.
Вылупившаяся
особь,
найдя
по
соседству
в
яйцевом
пакете
такое
яйцо,
прокусывает
его
оболочку
челюстями
и
постепенно
поглощает
содержимое.
Если
кормового
яйца
нет,
то
она
может
съесть
и обычное.
Муравьи
не
только
кормят
личинок,
но
и непрерывно
ухаживают
за
ними.
Облизывание
личинок
—
постоянное
и
самое
обычное
занятие
муравьев-нянек.
В
результате
тело
личинки
всегда
остается
чистым.
Кроме того,
рабо-
234
чие
регулярно
удаляют
экскреты
личинок
—
капельки
жидкости
из
анального
отверстия.
Интересно,
что
личинки
примитивных
муравьев
выделяют
эти
капельки
самопроизвольно,
а
рабочие
подбирают
их
позже.
У
более
высокоорганизованных
видов
личинки
выделяют
экскреты,
как
правило,
в
ответ
на
прикосновения
рабочих
к
заднему
концу
их
тела.
Муравьи
берут
эти
капельки
челюстями
и
либо
относят
на
«свалку»,
либо
выпивают.
Рабочие
удаляют
также
твердые
экскреты
личинок
—
мекониумы.
Не
менее
важной
формой
ухода
является
помощь
при
линьках.
Личинки
многих
высших
муравьев
нередко
без
помощи
нянек
не
могут
успешно
освободиться
от
сбрасываемой
шкурки
и даже
гибнут.
Столь
тесная
зависимость
личинок
от
ухаживающих
за ними
муравьев
приводит
к
неудачам
при
попытках
исследователей
вырастить
личинок
в
лабораторных
условиях
без
взрослых
муравьев.
Этим
пробовали
заниматься
многие,
но
с
незначительным
успехом.
Кормление
личинок
обычно
удается
наладить:
им
можно
подносить
на
кончике
иглы
прямо
ко
рту
кормовые
яйца,
кусочки
насекомых
или
грибницы
и
даже
поить
их
из
пипетки.
Но
муравьев,
облизывающих
личинок
и
удаляющих
их
экскреты,
не
заменить
никак.
Поэтому
вскоре
личинки
загрязняются,
на
их
теле
появляется
плесень
или
развиваются
другие
микроорганизмы,
и
личинки
гибнут.
Обычно
из
сотен
удается
довести до
окукливания
лишь
единицы!
Когда
выросшая
личинка
собирается
строить
кокон,
муравьи приносят
кусочки
древесины,
травинки
и
прочие
сухие
соринки
и
забрасывают
ими
личинку.
На
них,
как
на
каркасе,
личинка
натягивает
паутинные
нити
и
плетет
кокон.
Когда
кокон
готов,
рабочие
очищают
его
от
мусора
и
уносят.
Если
опоры
для
строительства
не
будет,
личинки окуклятся
без
коконов.
У
низших
муравьев
молодое
насекомое
появляется
из куколки
в
коконе,
прогрызает
его
и
выбирается
наружу
самостоятельно.
Высшие
муравьи
обыкновенно
разрывают
кокон
и
помогают
насекомому
выйти
из
куколки.
Если
коконы
удалить
из
гнезда,
то
лишь
некоторым
особям
удается
выйти
из
них
наружу,
а
большинство
гибнет
внутри
коконов
не
в
силах
избавиться
от
куколочных
шкурок.
У
муравьев,
личинки
которых
окукливаются
без
кокона,
помощь
при
выходе
из
куколки
нужна
не
в
меньшей
степени.
Только
у
некоторых
видов
(например,
у
мирмик)
части
рабочих
удается
самостоятельно
освободиться
от
куколочных
покровов,
у
многих
же
это
невозможно,
и
все
муравьи
погибают.
Особенно
нуждаются
в
содействии
при
выходе
из
куколки
длинноногие
муравьи
(афеногастер,
катаглифис
и
др.),
а
также крылатые
самки
и
самцы.
Факты,
приведенные
выше,
иллюстрируют
одну
из
важнейших тенденций
в
эволюции
общественных
насекомых
—
возрастание
взаимозависимости
особей
в
колонии.
Это
выражается
в усилении
и
усложнении
заботы
о
потомстве.
Возникают
235
и совершенствуются процессы переработки пищи, подготовки ее К поглощению личинками. В питании личинок все большую роль начинают играть кормовые яйца, секреты специальных желез. Зависимость развивающихся насекомых от взрослых собратьев становится полной и безусловной. В то же время и взрослые насекомые оказываются все более нужными друг другу. Взаимопомощь при питании и выполнении «требований гигиены» превращается из желательной в обязательную и жизненно необходимую.
СПОСОБЫ ОБЩЕНИЯ
Так муравьи, столкнувшись где-нибудь. А. Данте. Божественная комедия |
В
прошлом
мало
у
кого
возникали
сомнения в
том,
умеют
ли
муравьи
или
пчелы
разговаривать
друг
с
другом.
Это
казалось
очевидным.
Как
же
иначе
они
могли
бы
действовать
столь
разумно
и
согласованно?
Споры
шли
лишь
о
том,
общаются
ли
насекомые
с
помощью
звуков
или
же
их
язык
основан
на
иных
физических
явлениях.
Сэр
Дж.
Лаббок
в
70-х
годах
прошлого
столетия
затратил
массу
усилий,
чтобы
выяснить,
слышат
ли
пчелы,
осы
и
муравьи.
«Я
снова
и
снова,
—
писал
он,
—
испытывал
их
самыми
громкими
и
резкими
шумами,
какие
мог
только
производить,
употребляя
копеечную
дудку,
собачий
свисток,
скрипку,
равно
как
и
издавая
самые
пронзительные
и
ошеломляющие
звуки,
на
какие
только
способен
мой
голос;
но
все
это
не
производило
на
них
никакого
влияния.
В
то
же
время
я
тщательно
избегал
выводить
из
этого,
что
они
глухи,
хотя,
кажется,
что
их
род слуха
сильно
отличается
от
нашего».1
Теперь-то
мы
знаем,
что
муравьи
и
термиты совершенно
глухи
к
обычным
звукам,
являющимся
колебаниями
воздуха;
у
них
нет
специальных
органов
для
восприятия
этих
звуковых
волн.
Но
зато
все
насекомые
отлично
чувствуют
вибрацию
твердого
субстрата,
на
котором
сидят;
ее
воспринимают
так
называемые
подколенные
органы,
расположенные
на
голенях
всех
шести
ножек.
Поэтому
насекомые
«слышат»
ногами.
Известно,
что
звуковые
волны,
распространяющиеся
в
воздушной
среде,
в
основном
отражаются
от
твердых
предметов.
Вот
почему оказались
тщетными
все
усилия
Лаббока!
1 Лаббок Дж. Муравьи, пчелы и осы. Наблюдения над нравами общежительных перепончатокрылых. М., 1898. С. 202—203.
236
В
70-е
годы
нашего
столетия
Е.
К. Еськов
выяснил,
что
пчелы
и
осы
в
отличие
от
муравьев
и
термитов
способны
воспринимать
некоторые
звуки
из
воздушной
среды
специальными
волосками
(так
называемыми
фонорецепторами)
на
голове.
Однако
чтобы
общаться
с
помощью
звуков,
надо
уметь не
только
слышать,
но
и
издавать
их.
Огромное
большинство
насекомых,
в
том
числе
и общественных,
не
имеет
специальных
органов,
производящих
звуки.
Пчелы
и
осы
используют
для
этой
цели
крыловую
мускулатуру.
При
быстрых
сокращениях
мышц
хитиновые
части
наружного
скелета
груди
вибрируют,
издавая
жужжащие
звуки.
«Голосовой
диапазон»
пчел
и
ос
довольно
широк,
они
могут
жужжать
на
разный
лад
—
низко
или
высоко,
отрывисто
или
продолжительно.
Изучение
спектров
издаваемых
ими
звуков
позволило
установить,
что
осы
и
особенно
медоносные
пчелы
используют
в
общении
довольно
много
звуковых
сигналов,
хорошо
различающихся
по
физическим
параметрам.
Ниже
мы
рассмотрим
несколько
примеров
звуковой
коммуникации у этих
насекомых.
Некоторые
муравьи
и
термиты
способны
издавать
звуки трением
друг
о
друга
твердых
частей
наружного
скелета.
Звуковые
аппараты,
базирующиеся
на
этом
принципе,
называются
фрикционными.
У
большинства
муравьев
подсемейства
мирмицин
на
верхнем
заднем
крае
второго
членика
стебелька
есть
направленный
назад
и
вниз
заостренный
кантик
(или
«коготок»),
а
на
верхней
стороне
основания
брюшка
расположено
ребристое
поле
—
участок
кутикулы
с
мелкими
поперечными
морщинками.
Когда
брюшко
движется
относительно
стебелька
вверх
или
вниз,
«коготок»
царапает
по
этим
морщинкам
и
раздается
очень
слабое
стрекотание, или,
как
выражаются
энтомологи,
стридуляция.
Ее
можно
расслышать,
прижав
ухо
к
лабораторному
формикарию
с
возбужденными
мирмиками.
Стридуляционный
аппарат
мирмеций
и
понерин
устроен
примерно
так
же,
но
стрекочут
эти
муравьи
гораздо
громче
мирмик,
их
звуковые
сигналы
хорошо
слышны
с
расстояния
в
несколько
метров.
Нотомирмеция
стридулирует
значительно
тише,
фрикционный
аппарат
находится
у
нее
на
нижней
стороне
брюшка.
Некоторые
термиты
могут
издавать
скрипящие
звуки трением
головы
о
переднегрудь.
Но
все
же
множество
муравьев
и
термитов не
имеет
специальных
органов,
производящих
звуки.
Поэтому
они
просто
ударяют
о
субстрат
челюстями,
головой
или
брюшком.
Такой
стук,
распространяющийся
в
виде
колебаний
субстрата,
воспринимается
другими
насекомыми
на
порядочном расстоянии.
П.
Гюбер
был
первым
среди
ученых,
обратившим внимание
на
роль
усиков
в
общении
между
муравьями.
«Я
часто
видел,
—
писал
он,
—
как
они
пользовались
усиками
на
поле
битвы
с
целью
указать
на
приближение
опасности
и
отличить
себя
от
неприятеля;
среди муравейника,
чтобы
возвестить
о
появлении
237
солнца, имеющего такое благоприятное влияние на развитие личинок; во время
своих путешествий и передвижений, чтобы указывать друг другу дороги; отправляясь
на добычу корма; для определения времени начала похода и т. д.».2
В 1899 г. известный немецкий ученый, монах ордена иезуитов Э. Васманн, посвятивший
многие годы исследованиям муравьев и термитов, объявил, что ему удалось обнаружить
у них язык. Словами этого языка, по его мнению, являются прикосновения усиков-антенн
одного насекомого к телу, голове и антеннам другого. Васманн описал целый
ряд подобных сигналов-прикосновений, имеющих якобы вполне определенный смысл,
и утверждал, что муравьи и термиты способны передавать очень сложные сообщения,
эквивалентные фразам нашего языка. Концепция «антеннального языка», или «антеннального
кода», Васманна получила широкое распространение. Есть у нее сторонники и
в наше время. Насколько близка к истине эта точка зрения?
Сейчас прикосновения одного насекомого к другому антеннами, ножками и иными
частями тела, передающие какую-то информацию, принято называть тактильными (осязательными) сигналами. Ученые, пытающиеся их изучать, сталкиваются с
огромными трудностями. Движения насекомых очень быстры, и человеку порой
невозможно даже «схватить» их глазом, а тем более точно описать. Для этого
используют скоростную киносъемку, которой не было во времена Васманна. Но
мало подробно описать тактильные сигналы, надо выяснить их значение, если
таковое имеется. А как это сделать? Допустим, мы видим, что после того или
иного сигнала насекомое, которому он адресован, совершает определенное действие
и так происходит в большинстве случаев. Можно ли на этом основании утверждать,
что данный тактильный сигнал передает информацию о необходимости именно такого
действия? Конечно, нет! Мы же не исключили возможности одновременной сигнализации
с помощью запахов, звуков или каким-то иным путем. Например, Васманн интерпретировал
очень быстрые постукивания антеннами как сигнал тревоги. Но в его время еще
не было известно, что возбужденный муравей может выделить химическое вещество
со специфическим запахом — феромон тревоги, который и сигнализирует об опасности
прежде всего.
Как исследовать независимо роль химических и тактильных сигналов в этой ситуации?
Ведь тактильные сигналы тоже могут нести определенную информацию, дополняющую
химические сигналы. Но как ее выявить? Создать модель муравья, воспроизводящую
характерные прикосновения антенн, но лишенную феромонов? Это могло бы очень
приблизить нас к истине. Но пока никому не удалось подобное тончайшее моделирование
—
2 Huber P. Recherches sur les moeurs des fourmis indigenes. Paris; Geneve, 1810. P. 123.
238
слишком уж велики технические трудности. Правда, кое-что с моделями и муляжами
уже получилось, но на более крупных насекомых — пчелах и осах. И об этом
мы еще поговорим.
Все эти объективные трудности явились причиной того, что тактильная коммуникация
у общественных насекомых изучена пока хуже всех остальных форм общения.
Васманн и его последователи интерпретировали наблюдаемые ими тактильные
сигналы чисто
умозрительно, приписывая им то значение, которое, как они полагали, наиболее
соответствует ситуации. Понятно, что подобные построения сторонники строгих
и объективных методов экспериментальной науки встречали с недоверием.
До сих пор концепция антеннального языка в понимании Васманна и его единомышленников
не получила сколько-нибудь серьезного подтверждения. Из этого, конечно,
не следует, что тактильные сигналы не играют никакой роли в процессах
коммуникации
у общественных насекомых. Напротив, их значение в общении необычайно
велико.
Но прежде чем приводить доказательства этого утверждения, отметим, что
язык насекомых можно изучать, не только анализируя используемые при общении
сигналы.
Можно попытаться выяснить, что вообще способны «сказать» друг другу насекомые?
В состоянии ли они передать настолько сложную информацию, которой можно
обмениваться лишь при наличии языка? Ответить на этот вопрос проще, чем
стараться выяснить
смысл отдельных сигналов. Какие же результаты достигнуты на этом направлении?
Анализируя имеющиеся в науке данные, следует признать, что до настоящего
времени не получены строгие доказательства существования у общественных
насекомых языка, хотя бы в некоторой степени подобного человеческому.
Конечно, у общественных
насекомых есть довольно сложные формы передачи информации, но основаны
они на совершенно иных принципах, чем у людей.
Итак, тактильные сигналы вряд ли могут передавать сложные сообщения.
И далеко не всегда прикосновения усиками вообще выполняют сигнальную
функцию.
Насекомые
прикасаются друг к другу в первую очередь с целью ориентации, чтобы узнать
по запаху, кто находится перед ними — друг или враг, рабочий, солдат,
царица или личинка. Усики насекомое гораздо чаще использует именно для
получения
информации, а не для передачи. В каких же случаях доказано сигнальное
значение прикосновений?
Прежде всего без тактильной коммуникации невозможно взаимодействие насекомых
при трофаллаксисе. Понаблюдаем за тем, как рабочие рыжей мирмики кормят
личинок. Муравей-нянька заранее не знает, какие личинки, во множестве
лежащие в камере
гнезда, готовы принять от него корм. Он может определить это только путем
общения с каждой личинкой. Рабочий подходит к личинке и слегка постукивает
кончиками уси-
239
ков по ее голове и ротовым органам. Если личинка голодна, она отвечает
шевелением челюстей или движениями головы вверх-вниз. Уловив усиками
движения личинки,
муравей тут же отрыгивает ей в рот капельку пищи. Но личинка, которую
недавно накормили, не реагирует на прикосновения усиков рабочего.
Тогда он начинает
ее облизывать. Быстро движется язычок муравья, тщательно обрабатывая
голову и ротовые органы личинки. Время от времени рабочий прерывает
облизывание, постукивает усиками по голове личинки и опять принимается
ее облизывать.
В конце концов ему удается «растормошить» личинку и она подает муравью
сигнал, «соглашаясь» принять корм. Нередко можно наблюдать и обратную
ситуацию:
личинка
настолько голодна, что реагирует сильными движениями головы и челюстей
на случайные касания проходящих мимо муравьев и даже на сотрясение
гнезда. Муравьи
ощущают эти движения личинки, когда ощупывают усиками ее тело и вознаграждают
ее капелькой корма.
Таким образом, кормление личинки муравьем невозможно без довольно
сложного общения между ними, основанного на обмене тактильными сигналами.
Подобную
же картину можно видеть и у пчел-аллодапин. Ш. Сакагами наблюдал,
как личинки видов рода экзонеура движением челюстей и головы сигнализируют
о том, что
им нужен корм; в ответ на это пчелы захватывают их челюстями и отрыгивают
им в рот прозрачную жидкость.
Трофаллаксису между взрослыми насекомыми также предшествует обмен
тактильными сигналами. Для всех видов общественных насекомых характерны
специфические
стереотипные позы выпрашивания и предложения пищи, которые, вероятно,
тоже имеют сигнальное значение. Медоносная пчела выпрашивает корм
у другой особи,
всовывая хоботок между ее челюстями и одновременно постукивая усиками
по ее голове и ротовым органам. Поза предложения пищи выражается
в небольшом выдвижении хоботка вперед-вниз и отрыгивании капли корма.
Отделенная
от
тела голова вполне заменяет живую пчелу — другие рабочие будут предлагать
ей корм
и пытаться выпрашивать у нее. Голова без усиков гораздо менее эффективна.
Но если усики имитировать проволочками, то голова вновь станет объектом
предложения и выпрашивания корма. Некоторую роль в этом поведении
играет
и запах: небольшой
шарик из любого материала, лишенный даже подобия антенн, но обладающий
запахом пчелиной головы, иногда побуждает окружающих пчел выпрашивать
или предлагать
корм.
Тактильная коммуникация при трофаллаксисе у германской осы значительно
сложнее. Оса выпрашивает корм у предполагаемого донора, постукивая
усиками по его
голове и челюстям. Во время трофаллаксиса реципиент продолжает постукивание
— в противном случае донор прервет передачу корма. Поэтому
240
|
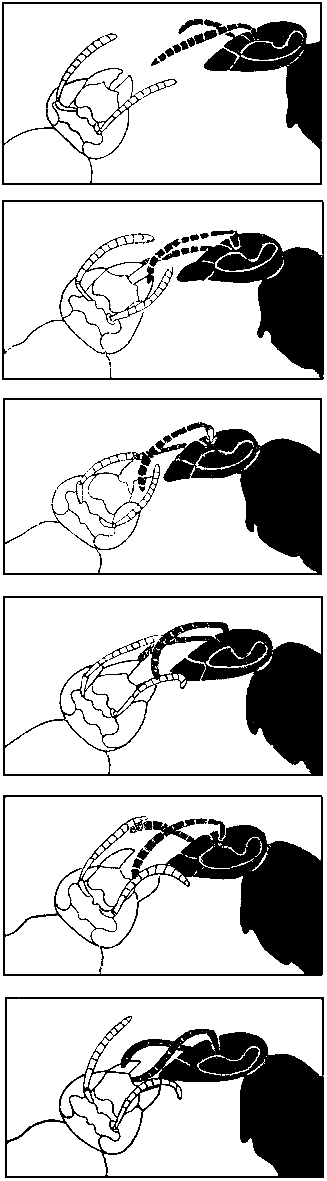
оса без антенн или даже лишенная их кончиков остается голодной. Отделенной
от туловища голове осы не предлагают пищу. Но если антенны заменить
проволочками, совершающими от 20 до 100 колебаний в секунду,
то такая модель головы
реципиента побуждает окружающих ос предлагать ей корм. Однако
уже через несколько
секунд оса-донор прерывает контакт и уходит, поскольку модель
не в состоянии правильно
воспроизвести прикосновения антенн к голове донора. Интересно,
что самцы ос не могут выпрашивать пищу у рабочих, видимо, потому, что
кончики их
усиков, загнутые крючком, совершенно непригодны для подачи соответствующих
тактильных
сигналов.
Лишенные антенн муравьи обычно тоже неспособны попросить пищу
у собратьев по гнезду. Выпрашивающий корм рабочий черного муравья-древоточца
поглаживает усиками голову и ротовые органы предполагаемого донора
и продолжает
делать это в течение всего трофаллаксиса. Усики его движутся
в
противофазе:
правый усик прикасается к левой стороне головы донора и скользит
по ней
до правой
мандибулы, затем левый усик проделывает тот же путь, но справа
налево. Донор вначале чаще всего вытягивает антенны над головой
реципиента,
а затем во
время передачи корма двигает ими довольно беспорядочно, но не
прикасается к реципиенту. Сходным образом ведут себя при тро-фаллаксисе
и муравьи
других видов: реципиент постукивает антеннами по голове и челюстям
донора, а тот
отводит антенны в стороны.
241
У многих видов предлагающий пищу муравей принимает характерную
позу. Рыжая мирмика, например, широко раскрывает челюсти
и замирает, удерживая
на язычке
отрыгнутую капельку корма. Сходным образом ведут себя и другие
муравьи. Нередко при этом поза муравья изменяется в зависимости
от того, сколь
сильно он «желает»
отдать корм. Рабочий красногрудого муравья-древоточца при
настойчивом предложении корма приоткрывает челюсти, поворачивает голову
примерно на 90° и приближает
ее к голове предполагаемого реципиента, одновременно поглаживая
его усиками. Если муравей, которому предлагают корм, не спешит
его взять,
донор сильно
наклоняется на бок, поворачивает голову еще на 90° и подставляет
ее под челюсти партнера. Обычно столь «настоятельная просьба»
приводит к желаемому
результату.
Насекомое, которому адресована сигнальная поза, воспринимает
ее зрением и осязанием, т. е. прикасаясь антеннами к партнеру.
Поэтому
такие
позы относят
к тактильно-зрительным сигналам. Но если дело происходит
в гнезде, где света нет, то тактильное восприятие становится
основным.
До сих пор мы
не знаем,
насколько зрительное восприятие важно в тех или иных случаях.
Количество известных сигнальных поз также невелико. Когда
муравьи
некоторых
видов рода лептоторакс переселяются в новое гнездо, многие
рабочие принимают
перед носильщиками
позу «предложения»: муравей поворачивается задним концом
тела и немного подгибает брюшко, выставляя членики стебелька,
за
который его и берет
носильщик.
Среди тактильно-зрительных сигналов гораздо более распространены
не позы, а сигнальные движения, совершаемые всем телом или
его частью и нередко
сопровождаемые прикосновениями усиками, ножками или другими
органами к окружающим насекомым.
Такие сигналы являются комплексными, их воспринимают тактильно,
по сотрясению субстрата, а при наличии света — зрением. Когда
муравей-носильщик рыжей
мирмики намеревается транспортировать рабочего в новое гнездо,
он
совершает
в его
сторону несколько коротких бросков, прикасаясь каждый раз
к приглашаемому муравью антеннами и даже слегка толкая его
челюстями.
Рабочий
обычно реагирует на это, подгибая ноги и антенны, а затем,
когда носильщик
захватывает его
за голову, окончательно принимает стереотипную позу переносимого
муравья. Разнообразные сигнальные движения широко используют
разные виды общественных
насекомых для мобилизации, предупреждения об опасности и
в других случаях. Очень сложными сигнальными движениями являются
так называемые
танцы
медоносных пчел. Обо всем этом мы еще поговорим специально.
Существуют и чисто зрительные сигналы, воспринимаемые на
значительном расстоянии. Но роль их исследована плохо. Пока
известен лишь
один достоверный случай
использования
242
зрительной коммуникации при мобилизации на пищу у муравьев. Пчелы-сторожа, дежурящие у входа в улей, хорошо отличают по полету и характеру движений пчел-сборщиц от других насекомых, а также от пчел-воровок, пытающихся проникнуть в улей за медом. Воровки обычно движутся несколько иначе, чем сборщицы, как-то неуверенно, что сразу же замечают сторожа. Но
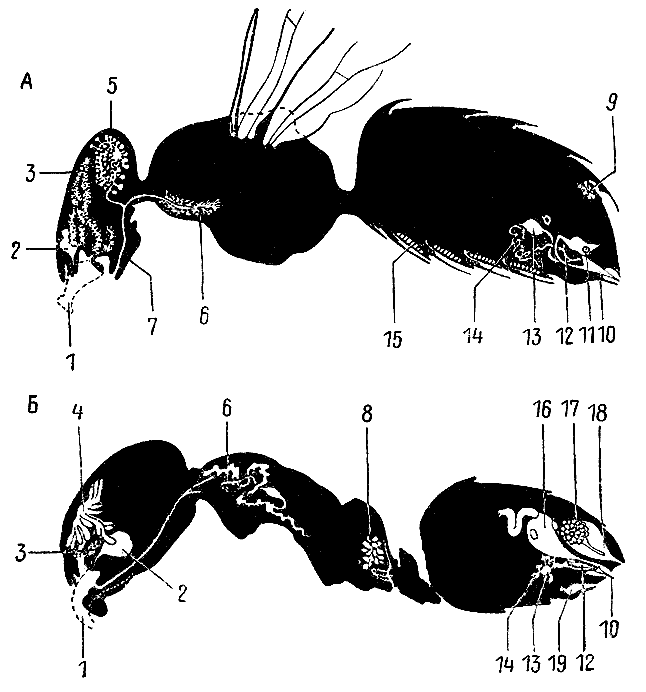
71. Железы внешней секреции (по Э. Вильсону, 1971 г.).
А — медоносная пчела; Б — муравей иридомирмекс хумилиса. 1 — мандибулы; 2 — мандибулярная железа; 3 — гипофарингеальная (максиллярная) железа; 4 — глоточная (фарингеальная) железа; 5 — головной отдел нижнегубной (лабиальной) железы; 6 — грудной отдел лабиальной железы (у муравья — вся железа); 7 — гипостомальная (защечная) железа: 8 — метаплевральная железа; 9 — железа Насонова; 10 — жало; 11 — железа Кожевникова; 12 — железа Дюфура; 13 — резервуар ядовитой железы; 14 — ядовитая железа; 15 — восковые железы; 16 — железы в задней кишке; 17, 18 — анальная железа и ее резервуар; 19 — железа Павана.
в данном случае вряд ли можно говорить о сигнализации
в истинном смысле слова.
Многие ученые считали, что характерные позы тревоги
и угрозы, принимаемые муравьями или медоносными
пчелами, окружающие
насекомые воспринимают
зрением и реагируют на них, как на сигнал опасности.
Но ведь одновременно насекомое
выделяет и феромон тревоги, являющийся сигналом
опасности. Сигнальная функция феромонов тревоги
очень хорошо
известна, а сигнальная
роль поз тревоги
и угрозы не доказана экспериментально. Так что
вопрос о чисто зрительном восприятии
сигнальных поз остается открытым.
243
Чрезвычайно широко распространена у общественных
насекомых химическая коммуникация с помощью
сигнальных веществ
— феромонов, выделяемых
во внешнюю среду специальными
железами. Таких желез у общественных насекомых
известно очень много.
Сигнализация посредством феромонов исследована,
пожалуй, лучше всех остальных форм общения
в связи с относительной
несложностью
и достоверностью
используемых
методов. Например, довольно просто при должной
сноровке выделить из тела насекомого ту или
иную железу и
приготовить ее экстракт
в эфире,
хлороформе,
гексане или ином органическом растворителе.
Такой экстракт можно затем нанести на любой
предмет
или испарить в
определенном объеме
воздуха
и выяснить реакцию
насекомых на содержащийся в нем феромон. На
помощь исследователям химической сигнализации
пришли
и тончайшие методы современной
физической химии
— газовая хроматография и массовая спектрометрия.
Они позволяют не только
определить
химическую природу феромонов, но и разделять
сложные смеси веществ, чтобы изучить роль их
компонентов.
Феромоны принято классифицировать на две группы.
К первой относятся феромоны-релизеры. Они воздействуют
на насекомое
через органы
обоняния, вызывая немедленный
ответ центральной нервной системы и изменения
в поведении насекомого. Большинство известных
феромонов
принадлежит
именно к этой группе.
Составляющие вторую группу феромоны-праймеры не вызывают немедленной реакции. В некоторых
случаях
насекомые
воспринимают их органами
обоняния, в других
праймеры попадают в организм с пищей и воздействуют
на эндокринную систему. Это обусловливает физиологические
изменения в организме,
что может отражаться
и на поведении насекомых. Таким образом, феромоны-праймеры
вызывают отсроченные реакции. К этой группе
относятся, например, феромоны
царской пары термитов,
регулирующие процессы развития насекомых разных
каст, царские феромоны общественных перепончатокрылых,
подавляющие плодовитость
рабочих
и препятствующие появлению
новых плодовитых самок. Интересно, что феромоны
цариц
служат одновременно и релизерами, поскольку
вызывают помимо отсроченных
эффектов и
непосредственные поведенческие реакции — стимулируют
фуражировочную и строительную
активность рабочих, влияют на их поведение
по отношению к личинкам и т. д.
По характеру действия на поведение насекомых
феромоны чрезвычайно разнообразны. Они участвуют
практически
во всех процессах
общения и передают всевозможные
сигналы. В дальнейшем мы познакомимся с основными
классами феромонов. Но несомненно, что первичной
функцией феромонов
в эволюции было
привлечение. Недаром у необщественных
насекомых факти-
244
чески единственными являются так называемые
половые феромоны, способствующие встречам
самцов и самок.
Феромоны, обеспечивающие
привлечение,
получили название аттрактантов.
Привлечение — это, несомненно, фундаментальная
форма коммуникации у общественных насекомых,
поскольку социальный образ жизни
немыслим без
стремления находиться
вместе, создавать скопления. Поместим небольшое
количество
муравьев или медоносных пчел в какой-нибудь
пустой закрытый сосуд. Сначала
встревоженные насекомые
беспорядочно рассеиваются в нем, блуждая
туда-сюда. Но постепенно они успокаиваются
и начинают
собираться в
тесные кучки. С
течением времени
они, скорее всего,
образуют одно скопление на дне сосуда.
Какие стимулы вызывают подобное поведение?
Чаще
всего это именно
феромоны-аттрактанты. Привлечение
участвует
в качестве
компонента и в других, более сложных системах
коммуникации — мобилизации, сигнализации
об опасности.
На поверхности тела общественных насекомых
всегда есть феромоны-аттрактанты. Достаточно
подробные
данные о
их химической природе и выделяющих
их железах получены пока лишь для немногих
видов. Так, в задней кишке желтошеего
термита при переваривании древесины образуется
несложное органическое вещество
З-гексен-1-ол. Оно распространяется по
всей поверхности тела и в значительной
степени обеспечивает
взаимную привлекательность термитов. Наибольшей
привлекательностью обладают личинки, наименьшей
— солдаты. Если термитов
кормить чистой целлюлозой
или лишить кишечных симбионтов, то привлекающий
запах постепенно исчезает. Вероятно,
аттрактант синтезируется в результате деятельности
жгутиконосцев из каких-то компонентов древесины,
отсутствующих в чистой
целлюлозе.
У большинства муравьев специальный аттрактант
выделяет железа Дюфура. Феромон железы
Дюфура муравьи используют
для привлечения
других
рабочих при мобилизации
и в некоторых иных ситуациях.
Между пятым и шестым сегментами на верхней
стороне брюшка медоносных пчел расположена
железа Насонова,
выделяющая
аттрактант с очень
приятным запахом.
Он представляет собой смесь, основными
компонентами которой являются гераниол,
гераниевая кислота
и цитраль. Пчелы
применяют этот
феромон в самых различных
ситуациях. Стоящие у входа в улей пчелы-вентилировщицы,
как правило, выпячивают железу Насонова.
Запах ее феромона насыщает
струю
воздуха, вытекающую из
летка, и помогает отыскивать улей возвращающимся
из полетов рабочим. При роении пчелы концентрируются
вокруг
матки,
привлекаемые запахом
ее феромонов.
Но этот запах они способны воспринимать
только на близком расстоянии. Вот почему
пчелы,
окружающие матку, выделяют
аттрактант из
железы Насонова, побуждающий всех остальных
пчел со-
245
браться вокруг матки плотным роем. Феромон
железы Насонова пчелы используют и
для мобилизации.
На нижней стороне лапок ос-веспин и
медоносных пчел обнаружены небольшие
железистые
образования — так
называемые железы
Арнхардта. Природа
выделяемого ими феромона неизвестна,
но установлено, что поверхность, по
которой передвигались пчелы и осы,
вызывает пристальное
внимание других особей того же вида.
Поэтому вход в гнездо пчелы и осы отыскивают
по знакомому запаху. Когда несколько
пчел побывает на кормушке с не имеющим
запаха
сахарным сиропом, то она становится
привлекательной для
других пчел, поскольку
их предшественники
оставили на
ней пахучие следы своих лапок.
Аттрактанты содержатся в так называемой
линочной жидкости, выделяющейся между
сбрасываемой шкуркой и телом термитов.
Именно эти вещества
привлекают грумов
и заставляют их облизывать линяющих
насекомых.
В некоторых случаях эффект привлечения
могут обеспечивать не специальные феромоны,
а иные
вещества. Например,
для многих муравьев очень
притягателен углекислый газ. Если пустить
в лабораторное гнездо
рыжей мирмики
слабую струйку углекислого газа, то
муравьи сразу же устремятся к точке,
откуда
она исходит.
В экспериментах Э. Вильсона с огненным
муравьем было установлено, что рабочие
этого вида
отлично умеют
ориентироваться по
градиенту концентрации
углекислого
газа, двигаясь обычно в направлении
ее увеличения. Это позволяет им присоединяться
к группам
муравьев, от которых
во все стороны
распространяется выделяющийся
при дыхании углекислый газ. Если несколько
муравьев засыпать тонким слоем песка,
то другие рабочие
начинают выкапывать
пленников. Установлено,
что землекопы ориентируются по запаху
углекислого газа, просачивающегося
сквозь
песок от зарытых муравьев. Запах углекислого
газа чувствуют медоносные пчелы и,
вероятно, многие
другие насекомые.
Феромоны, выделяемые царицей и царем,
прежде всего обеспечивают репродуктивный
контроль,
о чем уже
говорилось. Но одновременно
они являются аттрактантами
для рабочих. Синтетическая кетодеценовая
кислота — феромон царицы медоносных
пчел — позволяет
создать муляж, заменяющий
матку.
Если смочить раствором
этого вещества любой небольшой предмет
в улье, то пчелы
образуют вокруг него свиту и будут
вести себя совершенно так же, как
с настоящей царицей.
Легко
удается этот опыт и с синтезированным
химиками феромоном царицы восточного
шершня. Можно
получить активные
экстракты царских
феромонов муравьев,
термитов и безжалых пчел. Достаточно
положить физогастричную царицу черного
садового
муравья, например, в пузырек с эфиром,
чтобы через несколько дней проделать
эффектный опыт: любой
пористый предмет
(сердцевина веточки
бузины,
губка и т. п.), смоченный этим
246
экстрактом, будет долго «заставлять»
муравьев обращаться с ним, как
с царицей. Если царицу
муравья-кочевника
рода эцитон
подержать
некоторое
время на
кусочке дерева или картона, то
он станет крайне привлекательным для рабочих
муравьев.
Такой опыт удается и с другими
муравьями, в частности с кампонотусами. Вот
насколько сильным
аттрактантом
является феромон царицы!
Царские феромоны способны очень
долго сохраняться. В гнезде огненного
муравья
труп царицы
не утратил привлекательности для
рабочих девять недель. А рабочие
муравья-портного продолжали «ухаживать»
за высохшим трупом царицы полгода.
Но рекорд
здесь принадлежит,
видимо, медоносной
пчеле:
сухая матка, свыше десяти
лет находившаяся на булавке в коллекционном
ящике, оказалась все еще в некоторой
степени привлекательной для пчел!
У царицы муравьев-портных на верхней
стороне брюшка Э. Вильсон и Б.
Хеллдоблер обнаружили
железы, открывающиеся
отверстиями
на небольших
выростах перепонки
между сегментами. Вот эти выросты
и привлекают особенно рабочих,
облизывающих их и все тело царицы.
У огненного
муравья
царица, видимо, выделяет аттрактант
из
ядовитой железы.
На верхней
стороне
брюшка находятся
продуцирующие
аттрактант железы у царицы безжалой
пчелы паратригона субнуда. Царица
очень часто
вибрирует крыльями и потирает ножками
брюшко, этим она способствует распространению
по
телу феромона.
Неодинаковое количество выделяемого
аттрактанта может лежать в основе
предпочтения рабочими
одной из нескольких
цариц.
Эксперименты с огненным
муравьем позволили
установить, что царицы значительно
различаются по привлекательности,
а рабочие чаще
всего предпочитают царицу, выделяющую
больше феромонов, нередко
убивая
остальных. Как показывают некоторые
данные, количество выделяемого
царицей аттрактанта
напрямую связано
с ее плодовитостью. Поэтому выбор
рабочими самой привлекательной
царицы вполне
оправдан. Различия в
привлекательности цариц
определяют очень интересный процесс
смены царицы, в деталях прослеженный
у вида
паратригона
субнуда.
У
этой безжалой
пчелы вышедшие из
куколок
плодовитые самки прячутся в районе
горшков с провизией и даже занимаются
там разнообразной
строительной деятельностью. Постепенно
молодая «претендентка
на престол» становится все более
привлекательной для пчел. Она выделяет
изо рта
капельки секрета мандибулярных
желез, которые поглощают
рабочие. Пчелы поедают
и экскременты царицы. Вероятно,
в мандибулярном
секрете и экскрементах тоже
содержатся
феромоны, но это пока не исследовано.
Брюшко молодой царицы увеличивается,
и вокруг
нее появляется
обширная
свита пчел,
которые облизывают
царицу, особенно кончик ее брюшка,
и вступают с ней в трофаллаксис
— в процессе
его царица
247
издает характерный запах, хорошо ощутимый даже человеком. В это время молодая
царица либо покидает гнездо и с роем пчел переселяется в новое, либо
пытается сместить старую царицу.
В последнем случае молодая царица в сопровождении свиты приближается
к старой и, возбужденно вибрируя крыльями, наносит на ее брюшко капельку
мандибулярного
секрета, а иногда еще и капельку из анального отверстия. Старая царица
начинает отчищаться, ее облизывают и окружающие пчелы. Если старая царица
еще плодовита,
она, видимо, выделяет достаточно феромонов и сохраняет привлекательность
для рабочих. В этом случае пчелы не позволяют молодой царице «обижать»
старую, прогоняют и даже убивают ее, если она не может покинуть гнездо
с роем. Но
если феромоны у молодой царицы оказываются «сильнее», чем у старой, то
она еще и еще раз пачкает старую царицу своим секретом. В конце концов
это вызывает
у пчел враждебность к старой царице, они нападают на нее и раздирают верхнюю
сторону брюшка, где находятся кожные железы. Молодая царица становится
единственной в гнезде. «Дворцовый переворот» завершился!
Есть у общественных насекомых и половые феромоны-аттрактанты. Трутни находят
в воздухе летящую матку по сохраняющемуся запаху ее кетодеценовой кислоты,
которая, таким образом, выполняет функции и полового аттрактанта. Самки
и самцы термитов оставляют за собой пахучие следы с помощью той же железы,
что выделяет следовой феромон у рабочих термитов (подробнее см. в подразделе
«Мобилизация»). Самцы шмелей производят половой аттрактант в мандибулярных
железах. Крылатые самки некоторых муравьев, в особенности эргатоидные царицы
ряда видов (харпагоксенусы и др.), привлекают самцов, принимая призывную
позу с поднятым кверху брюшком и выделяя аттрактант. Самцы муравьев-древоточцев,
вылетая из гнезда, выделяют какой-то феромон, побуждающий самок присоединиться
к их полету.
В очень редких случаях общественные насекомые используют для привлечения
себе подобных не феромоны, а звуковые сигналы. Еще Э. Рубо установил, что
«вибрация крыльев осы-няньки служит сигналом для личинки, которая для того
чтобы получить корм, высовывает голову из ячейки... Достаточно бывает издать
громкий свист или другой резкий звук возле гнезда белоногастера — и головы
всех личинок показываются из отверстий».3 Голодные личинки
ос-веспин, двигая головой, скребут челюстями по стенкам ячеек, издавая
характерный «шуршащий»
звук. Личинка подает сигнал каждые несколько секунд. Осы воспринимают возникающую
при этом вибрацию сота,
3 Roubaud E. Recherches biologiques sur les guepes solitaires et sociales d'Afrique. Paris, 1916. P. 95—96.
248
быстро находят голодных личинок и кормят их. Интересно, что если таких личинок в гнезде несколько, то они скребут челюстями синхронно. Быть может, призывные сигналы в «хоровом исполнении» быстрее доходят до рабочих ос?
ВИЗИТНЫЕ КАРТОЧКИ
Этими наблюдениями, мне думается, установлен тот факт, что муравьи узнают не личность, или индивидуальность, каждого и что их согласие основано не на личном знакомстве каждого муравья со всеми остальными членами общины. Дж. Лаббок. Муравьи, пчелы и осы |
Колонии большинства общественных насекомых — это замкнутые системы,
вход в которые посторонним невозможен или, по крайней мере, затруднен.
Осы,
пчелы, муравьи и термиты обыкновенно не допускают в свои гнезда
чужаков не только
других, но и собственного вида, яростно набрасываясь на них,
прогоняя, а нередко и убивая. Такая замкнутость колонии свойственна
даже
примитивным эусоциальным видам, например галиктам, у входа в
норку которых всегда
дежурит пчела-сторож, пропускающая внутрь только своих, пришлых
же встречают разинутые
челюсти сторожа или высунутое из норки брюшко с обнаженным жалом.
Уже давно ученые пришли к выводу, что насекомые различают своих
и чужих по запаху. Они агрессивно реагируют на мертвых или усыпленных
посторонних,
которые
не могут издавать никаких сигналов, кроме химических, и одинаково
хорошо узнают находящихся в таком же состоянии насекомых из своего
гнезда
на свету и в темноте, следовательно, зрение не участвует в опознавании.
После «купания»
в органических растворителях, смывающих с тела запахи, чужаки
вызывают гораздо меньшую враждебность и нередко их принимают
миролюбиво.
Напротив,
насекомых,
обмазанных экстрактом чужаков, встречает в собственном гнезде
выраженная агрессивность. Подобные опыты, проводившиеся многими
исследователями
с самыми разнообразными видами общественных насекомых, привели
к заключению о наличии
специфического «общего запаха» семьи или колонии.
Чем же обусловлен этот общий запах? Вначале многие исследователи
были уверены, что все дело в экзогенных (внешних) источниках.
Общий запах
гнезда, согласно
их представлениям, формируется из запахов гнездового материала,
почвы и доставляемой в гнездо пищи; обитающие в гнезде насекомые
несут
его на поверхности
тела,
что и позволяет им легко узнавать
249
друг друга. Эксперименты подтверждали эту гипотезу. Если семью
пчел расселить в два улья, поставив их рядом, то сборщицы
будут постоянно
путать их,
залетая то в один, то во второй. Но когда пчел в одном улье
накормили сиропом с
запахом лаванды, а во втором — мятным сиропом, неразбериха
прекратилась и пчелы стали
отлично узнавать насекомых из своего улья и враждебно относиться
к соседям. И наоборот, когда пчел разных семей кормили сиропом
с одинаковым
сильным
запахом, агрессивность между ними уменьшалась, а иногда и
вообще исчезала. Если пчелы длительное время собирают нектар на определенном
растении,
например вереске, они начинают путать ульи, стоящие слишком
близко. Специальные опыты позволили установить, что кутикула пчел хорошо
поглощает запах
пищи.
Когда
нескольких шмелей посадили в клеточку и поместили ее на 2—3
часа
в другое гнездо, то после этого в собственном гнезде их встретили
весьма
недружелюбно.
Значит, на теле шмелей был запах чужого гнезда. Аналогичные
эксперименты с тем же результатом были проведены и на многих муравьях.
Рабочих
малого лесного муравья из одного гнезда поселили в два формикария,
изготовленных
из разных материалов, и в течение нескольких месяцев кормили
неодинаковой пищей. В результате оказалось, что муравьи из
этих формикариев
относятся друг к другу довольно враждебно.
Однако постепенно в науке накапливались данные о том, что
у общественных насекомых существуют эндогенные (внутренние)
источники
специфического
запаха колонии. Две семьи бурого лесного муравья разделили
каждую пополам и содержали
семь месяцев, как и в только что описанном опыте с малым
лесным муравьем, в формикариях из разного материала и на
различных
диетах. После этого
выяснилось, что муравьи проявляли некоторую агрессивность
к рабочим из своей семьи,
жившим в иных условиях, и несравнимо большую враждебность
к рабочим из чужой семьи,
обитавшим весь срок в точно таком же формикарии и питавшимся
той же провизией. Значит, внутренние различия запахов в данном
случае
оказались
важнее
приобретенных.
Хорошо известно, что муравьи из моногинных семей, как правило,
гораздо нетерпимее к посторонним, чем в обширных полигинных
колониях. Логично
предположить, что наследственно обусловленный индивидуальный
запах насекомых в семье
не
столь изменчив, как в генетически менее однородной полигинной
колонии. Поэтому по запаху моногинные колонии различаются
отчетливей, чем
полигинные.
Здесь нужно подчеркнуть одно обстоятельство: отсутствие выраженной
агрессивности еще не означает, что насекомые расценивают
запах чужака в качестве своего.
Как теперь стало ясно, открытая враждебность проявляется
только в случае явных различий между запахами чужака и хозяев
гнезда.
Когда
250
различия меньше, насекомые ощущают их и ведут себя с посторонними
без агрессивности, но иначе, чем с собратьями по гнезду.
Проиллюстрируем это примером. В природных
популяциях полигинных мирмик генетические различия обычно
возрастают с увеличением расстояния между гнездами. Если
мы пустим в гнездо
мирмик
рабочих,
пойманных
в нескольких километрах от данного места, то хозяева
прогонят или убьют их. Если взять таких же рабочих на дистанции
в десятки метров,
то их
примут весьма
настороженно, окружив со всех сторон и возбужденно ощупывая
усиками, и долго не позволят им ходить по гнезду. Ну,
а муравьи из соседнего
гнезда, отделенного
промежутком в 1—2 м, будут встречены без всякой видимой
реакции. Но когда мы внимательно присмотримся к поведению муравьев
и посчитаем количество
контактов между ними, то обнаружим, что хозяева гнезда
«игнорируют» чужаков: почти
не облизывают их и гораздо реже вступают с ними в трофаллаксис,
чем с собратьями
по гнезду. И так они будут вести себя до тех пор, пока
не привыкнут к запаху чужаков.
Наблюдения за поведением насекомых при общении их друг
с другом позволили установить, что наследственные различия
запаха есть
у всех изученных
в этом отношении видов. Более того, впервые была доказана
возможность индивидуального
узнавания насекомых по запаху.
В лабораторных экспериментах американскими исследователями
под руководством Ч. Миченера создавались искусственные
колонии из
куколок галикта
диаликтус зефирус, вырытых из природных гнезд. Оказалось,
что пчела-сторож запоминает
каждую особь в группе и пропускает в гнездо только знакомых
ей. Когда пчелу помещали на 2—3 дня в чужое гнездо или
наносили на
нее вещество
с посторонним
запахом, то сторож все равно принимал ее за свою. Но
если пчела отсутствовала 8—10 дней, сторож забывал ее
запах.
В первые
сутки после выхода из
куколки пчелы еще не имеют характерного запаха и их могут
принять в любое гнездо;
вскоре запах появляется, даже если пчелу содержать в
пробирке. Все это говорит о том, что узнавание своих
и чужих основано
у данного вида исключительно
на наследственных различиях запаха каждого насекомого.
Как было установлено
в дальнейшем, самцы этого галикта при спаривании с самками
также способны индивидуально различать их по запаху.
В экспериментах киевского энтомолога П. Г. Москаленко
с медоносными пчелами, когда их приучали находить кормушку
с сиропом по
запаху матки, удалось
доказать, что пчелы хорошо отличают запахи разных маток,
но запахи маток-сестер иногда
путают. Значит, царица медоносной пчелы обладает индивидуальным
запахом. Аналогичное явление было недавно обнаружено
и
для цариц рыжей мирмики.
Наблюдения за муравьями в небольших группах также позволяют
утверждать,
251
что они «знают друг друга в лицо» и ведут себя по-разному
в зависимости от того, с кем имеют дело.
Все эти для многих совершенно неожиданные данные
привели к подлинному перевороту в представлениях
о социальной
организации насекомых.
Они окончательно убеждают
нас в том, что общество насекомых не является «анонимным».
Муравьи, пчелы, осы способны узнавать друг друга
по запаху, если они «знакомы».
До недавнего
времени ученые были уверены, что индивидуальная специфичность
запаха присуща человеку и млекопитающим, может быть,
птицам и иным высшим
позвоночным, но никак не насекомым. Теперь ясно,
что это далеко
не так.
В последние годы была обнаружена еще более поразительная
способность насекомых различать по запаху степень
генетической родственности.
Оказалось, например,
что пчела-сторож галикта диаликтус зефирус, встречаясь
у входа в лабораторное гнездо с незнакомой пчелой,
в среднем тем чаще
пропускает ее в гнездо,
чем теснее их родственные отношения. Почти всегда
впускает она родных
сестер, реже — полусестер по отцу, еще реже — двоюродных
сестер и племянниц, еще
реже — полусестер по матери4 и т. д.
Самки полистов при закладке гнезда в лабораторных
экспериментах явно предпочитают объединяться с сестрами,
отвергая
попытки неродственных ос примкнуть к
их группе. Они отличают сестер от более дальних родичей,
даже если никогда не
встречались с ними в жизни. Когда самкам-основательницам
вместо собственного
гнезда предлагали чужое, они почти всегда принимали
и продолжали воспитывать расплод своих сестер, но
уничтожали неродственные
яйца и личинок; в
гнезде чужой самки они оставляли лишь куколок и несколько
крупных личинок, откладывая
в освободившиеся ячейки собственные яйца.
Подобные данные получены также для некоторых видов
муравьев и медоносной пчелы. Муравьи рода кампонотус,
например,
отличают неродственных
им рабочих, даже если они вышли из куколок в том
же гнезде (в
этих экспериментах
куколок переносили из одной семьи в другую): они
реже кормят и облизывают их по
сравнению с рабочими-сестрами. Муравьи менее усердно
ухаживают за личинками, если те
принадлежат к посторонней семье.
Надо добавить, что до самого последнего времени о
способности животных оценивать степень своей генетической
близости
с прочими индивидами
говорили только
в форме предположения. И нельзя не удивляться тому,
что эта способность впервые была обнаружена и изучена
именно
у насекомых,
а не среди
высших
4 У насекомых отряда перепончатокрылых коэффициент генетической родственности самки составляет: с сестрами — 3/4, с полусестрами, имеющими одного отца, — 1/2, с двоюродными сестрами и племянницами — 3/8, с полусестрами, имеющими одну мать, — 1/4.
252
позвоночных. Возможность различать степень родственности,
по-видимому, играет немалую роль в эволюции
социального поведения.
Где же источник индивидуального запаха насекомых?
Для ответа на этот вопрос пока мало данных.
Ясно лишь,
что запах создают
вещества,
находящиеся
на
поверхности тела. Для индивидуальной специфичности
запаха таких веществ должно быть достаточно
много. Тогда отдельные насекомые могли бы отличаться
по составу пахучей смеси и по количеству компонентов
в ней.
Эти предположения
удалось
подтвердить в последние годы. В составе кутикулы
муравьев родов кампонотус и одонтомахус
методом газовой хроматографии было обнаружено
множество углеводородов и других
органических веществ, которые могут быть источником
индивидуального запаха насекомого. Эти соединения
плохо испаряются, а
значит насекомые могут
воспринимать их только при близком контакте;
именно так и осуществляется, как мы знаем,
процесс узнавания.
Удалось обнаружить и отчетливые различия в
количественном составе смеси кутикулярных углеводородов
не только
между насекомыми
из разных семей,
но и между отдельными
муравьями. При этом, как и следовало ожидать,
муравьи из одной семьи различаются по углеводородам
очень
незначительно, а из
разных — довольно
существенно.
Смеси кутикулярных углеводородов генетически
близких семей
оказались более сходными, чем у менее родственных
и враждующих семей. Но
какие железы выделяют
кутикулярные углеводороды? Это пока неизвестно.
В последнее время выяснилось, что и некоторые
другие феромоны общественных насекомых могут
быть специфичными
для семьи
и даже для отдельных
особей. В экспериментах П. Г. Москаленко по
обучению медоносных пчел на запах
удалось обнаружить не только индивидуальную
специфичность феромона матки, но и
различия между семьями по запаху следовых веществ,
выделяемых железами Арнхардта на
лапках. Рабочие муравья-понерины пахикондила
тессеринода находят собственный запаховый след
среди множества
следов рабочих того
же гнезда. Рабочие
видов парапонера клавата и лептоторакс аффинис
тоже различают следы своих собратьев.
Значит, и следовые феромоны могут обладать
индивидуальной специфичностью. У многих видов
муравьев найдены
специальные феромоны, используемые
для мечения территории; их запах неодинаков
у разных семей и колоний одного
вида.
Итак, с современной точки зрения наследственные,
т. е. эндогенные, различия запаха существуют
у всех видов
общественных
насекомых
и всегда участвуют
в процессах узнавания. Экзогенные запахи играют
определенную роль, но далеко не у всех видов.
Например, у большинства
муравьев их
значение относительно
невелико, а у медоносной пчелы — весьма важно.
С пред-
253
ставлением о том, что запах семьи медоносных
пчел имеет целиком экзогенную природу,
приходится расстаться.
Точными экспериментами
доказано,
что даже в совершенно идентичных условиях
содержания и кормления пчелы
сохраняют способность к различению особей
из своей и чужих семей; они не проявляют
агрессивности
к чужакам и в то же время не путают их
со своими.
Но как быть с «общим запахом гнезда»? Существует
ли он? В свете современных знаний гипотеза
о том, что
запахи
отдельных насекомых
смешиваются
и образуют некий общий усредненный запах,
не подтверждается. Трудно представить,
как могут смешиваться нелетучие кутикулярные
углеводороды разных насекомых. Если даже
это в какой-то степени
и происходит, то
все равно невозможно
образование
их однородной смеси. Скорее всего, насекомое
оценивает меру
отличия запаха
встреченной им особи от как бы «эталонного»
запаха знакомых ему собратьев по гнезду.
Это не исключает тем не менее возможной
роли отдельных общих запахов, например
запаха
пищи, в процессе
узнавания, что
и показано для
медоносной пчелы.
Существует интересная гипотеза о том, что
феромон царицы, попадая на тела всех рабочих,
может обеспечивать узнавание ими друг друга.
Некоторые факты позволяют думать, что подобный
механизм играет
определенную роль у медоносной
пчелы. На теле
всех пчел есть феромон матки, который,
как мы знаем, обладает индивидуальной специфичностью
и поэтому
может участвовать
в определении их семейной
принадлежности.
Проведенные в последнее время эксперименты
с кампонотусами позволили подтвердить эту
гипотезу. Оказалось,
что кампонотусы враждебно
относятся даже к сестрам,
если те живут в гнезде с чужой царицей,
но лояльны
к неродственным рабочим, обитающим в гнезде
с их царицей. Следовательно,
специфичность запаха
муравья не зависит от его происхождения
и определяется только царицей. Но когда
царица имеет пониженную плодовитость и,
вероятно,
выделяет недостаточно феромона,
в узнавании главенствующую роль начинают
играть собственные запахи рабочих.
Не исключено также, что источником каких-то
запахов у рабочих могут быть и ароматические
вещества
собратьев. Это установлено,
например,
для тех
же кампонотусов. В экспериментах рабочим-нянькам
давали коконы из другой семьи,
и они выводили из них рабочих. Через две-три
недели этих
муравьев поселяли вместе со старшими рабочими
и наблюдали за их взаимоотношениями.
Старшие
рабочие, взятые из чужих семей, всегда
агрессивно реагировали на неродственных
молодых муравьев,
но их враждебность
проявлялась значительно меньше,
если они были сестрами нянек, воспитывавших
молодых рабочих. Аналогичные
результаты
получены и в сходных экспериментах с мирмиками.
254
Но вот у псевдомирмекс ферругинеа различия
запаха рабочих не зависят ни от присутствия
в гнезде
той или иной
царицы, ни
от того, кто
выкармливал этих
муравьев; они определяются феромонами,
выделяемыми самими муравьями. Следует
полагать, что виды общественных насекомых
значительно различаются по роли общих
приобретаемых запахов
в процессах узнавания
своих и чужих.
Откуда же насекомое знает, какой именно
запах присущ его семье, колонии, обладает
ли оно
«врожденным эталоном» запаха?
Еще
в 70-х годах XIX
в. О. Форелю удалось создать смешанные
группы муравьев,
поместив в формикарий
коконы разных видов рода формика. Выйдя
из куколок в одном гнезде, муравьи
вели себя в дальнейшем очень дружелюбно.
Американская
исследовательница муравьев А. Фильд
в начале XX в. сумела таким способом
получить группы даже столь
далеких родов, как формика, афеногастер
и
амблиопоне. В дальнейшем выяснилось,
что можно смешивать
не только куколок,
но и едва
вышедших из куколок
рабочих
— они еще легко уживаются друг с другом.
Точно так же создавали смешанные группы
разных видов
ос, шмелей,
безжалых
и медоносных
пчел.
Иногда удается получить сборные группы
муравьев и в более старшем возрасте.
Когда смешивали
крематогастеров из
разных семей одного
вида, между
ними начиналось настоящее сражение,
уносившее множество муравьиных жизней,
но постепенно
рабочие утихомиривались, драки прекращались,
и в
дальнейшем
насекомые жили без всякой враждебности.
Подобное выходит со многими муравьями.
Сложнее
«помирить» муравьев разных видов. В
одном
опыте соединили искусственные формикарий
пахучего
муравья-древоточца и обыкновенного
лесного муравья. Насекомые вначале
вели ожесточенные
схватки,
но позже «подружились»
настолько, что
сообща занимались
строительством, облизывали и кормили
друг друга. Однако древоточцы в конце
концов
все-таки убили
царицу лесных
муравьев.
Все эти эксперименты позволяют прийти
к выводу: насекомые могут привыкать
к чужому
запаху
и «признавать» имеющих
его особей
за «своих». Но
это, конечно, не означает, что они
не отличают видовых запахов. Как показали
наблюдения
за поведением групп муравьев, например,
родов кампонотус и формика, лазиус
и
крематогастер,
созданных методом
смешивания куколок,
муравьи, не проявляя
враждебности к рабочим другого рода,
все же явно предпочитают общаться с
особями своего
вида.
В общем, чем более
удалены по систематическому
положению виды муравьев, тем меньше
они общаются в смешанных группах. Не
исключено,
однако, что муравьи разных родов избегают
друг
друга еще и
вследствие различия тактильных сигналов.
Теперь уже хорошо доказано, что общественные
насекомые запоминают запах родного
гнезда, так сказать, в
самой ранней юности
— в первые дни после
выхода из
куколки. Именно в это
255
время у них осуществляется «запечатление»
(импринтинг) того или иного запаха,
который в дальнейшем
становится для них
«эталонным». Если
рабочим волосистого
лесного муравья в первые дни после
выхода из куколок дать коконы обыкновенного
лесного муравья,
то впоследствии
они
будут отвергать
и даже уничтожать
коконы своего вида, предпочитая
ухаживать за коконами обыкновенного лесного
муравья. В других опытах волосистым лесным муравьям
помогали выбраться из коконов и
ухаживали за
ними в первые
дни жизни няньки
обыкновенного лесного
муравья. И оказалось, что когда
эти волосистые
лесные муравьи повзрослели,
они
проявляли несомненную враждебность
к рабочим своего вида из того
же гнезда, откуда
были взяты куколки, но дружелюбно
относились к обыкновенным лесным муравьям.
Не исключено, что запоминание запаха
родного гнезда может происходить
еще на стадии
личинки. В экспериментах
с
черным бегунком яйца
из одной семьи
перенесли в другую, где из них
развились личинки. Когда личинки окуклились,
коконы вернули в
родное гнездо
и там из них вышли
муравьи. Затем
этих рабочих изъяли из гнезда и
дали им личинок из обеих семей. И оказалось,
что муравьи
усерднее ухаживали не за личинками
из собственной
семьи, т. е. родными сестрами,
а за личинками из гнезда, в
котором были
выкормлены,
т.
е. за «сводными сестрами».
Но через 15—20 дней рабочие утрачивали
«пристрастность» и уделяли одинаковое
внимание родным и сводным
сестрам. Если
муравьев
выращивали в чужой
семье не с яйца, а со стадии крупной
личинки, то они в дальнейшем предпочитали
ухаживать
за личинками-сестрами.
Значит, запах
родного гнезда бегунки
могут запоминать, еще будучи крохотными
личинками? Чтобы ответить на этот
вопрос, нужны дополнительные исследования.
Ведь экспериментаторы
не проверяли реакции «приемышей»
на
взрослых муравьев из
своей
и чужой семей.
Общественные насекомые не только
умеют отличать своих от чужих,
но и знают,
«кто есть кто»
в собственном гнезде. Царицу и
царя они узнают,
как уже
отмечалось, по их феромонам-аттрактантам.
Вероятно, любое плодовитое насекомое
обладает иным запахом, чем неспособное
к яйцекладке.
Царица полистов проявляет открытую
враждебность к другим
осам, пытающимся
отложить яйца.
При этом
ее особенно
привлекает кончик брюшка таких
самок — наверное, там и
выделяется специфический «феромон
плодовитости». Сходную картину
можно видеть у шмелей, ос-веспин.
Пчелы-трутовки выделяют немного
царского
феромона,
что и позволяет другим пчелам их
узнавать.
Плодовитые рабочие
отличаются
от
бесплодных по запаху
также у муравьев и, вероятно, у
всех общественных насекомых. Пожалуй,
по запаху насекомые
могут распознавать все
морфологические касты
и возрастные группы. Однако эти
вопросы
пока исследованы совсем плохо.
256
Муравьи хорошо различают возраст
личинок, скорее всего, по количеству
и длине
волосков на их
теле, но не исключено,
что и по запаху.
Нередко личинки
в
муравейнике рассортированы
как по возрастам, так и по размерам
в пределах
одного
возраста. Никогда
не
путают
муравьи кормовые
и нормальные
яйца,
даже если они близки по размерам.
А рабочие фараонова муравья
отличают, кроме
того, яйца, из которых появятся
самцы (в них одинарный, гаплоидный,
набор хромосом), от яиц, развивающихся
в самок (с двойным, диплоидным,
набором
хромосом). Распознают муравьи
и личинок, и куколок самок
и самцов.
Вообще же основным фактором,
позволяющим муравьям узнавать
расплод, является
не его внешний вид,
а запах специального
феромона, находящегося
на теле
личинок и куколок. Его муравьи
могут ощущать только на близком
расстоянии.
Если личинок
или куколок погрузить в гексан
или иной органический растворитель,
то
мы получим
экстракт феромона
расплода. Теперь можно
намочить им кусочки
дерева,
картона и прочие мелкие предметы
и дать их муравьям. И выяснится,
что
эти кусочки
не
менее привлекательны,
чем
настоящие личинки
и куколки:
муравьи
унесут их в гнездо и будут
держать вместе с расплодом, облизывать
и перетаскивать, как
и личинок, до
тех пор, пока не выветрится
запах феромона. Значит,
феромон расплода не только
служит для узнавания, но и стимулирует
поведение заботы
о потомстве.
В последние годы установлено,
что запах личинок и куколок,
развивающихся
в
крылатых самок,
отличается от запаха
личинок и куколок рабочих.
Рабочие мирмик, по данным М.
Брайена, весной легко узнают
личинок будущих
крылатых самок по запаху, который
наиболее силен
в области нижней стороны их
груди. Способны к этому и
муравьи других
родов. Рабочие
шмели тоже
хорошо отличают
личинок будущих самок-основательниц
от личинок
рабочих
и самцов. Вероятно, многие,
если не все, общественные насекомые
могут распознавать касты
еще на стадии личинки.
Химические стимулы позволяют
муравьям отличать мертвых насекомых
от живых.
Когда кто-нибудь
из собратьев
умирает внутри гнезда,
то рабочие
одних
видов скармливают его личинкам,
а других — выносят на специальные
«свалки»,
т. е. в места скопления гнездовых
отбросов.
Муравьи выбрасывают трупы только
после того, как в
них начинаются процессы
гниения и образуются
специфически
пахнущие вещества. Но если
для нас неприятный
трупный запах обусловлен в
основном аминами, индолами,
меркаптанами, то муравьи на
эти
вещества не реагируют.
Э. Вильсон с коллегами обнаружил,
что для них признаком гниения
является присутствие жирных
кислот с длинными
углеводородными
цепями (например,
олеиновой) и их эфиров. Если
удалить эти
вещества из трупов специальными
257
растворителями, то такие трупы муравьи оставят в гнезде. В то же время любой предмет, смоченный трупной вытяжкой, муравьи вынесут на свалку. Когда живых муравьев смазали этой вытяжкой и выпустили в гнездо, рабочие схватили их и, невзирая на попытки освободиться, отнесли на свалку «живые трупы». Заживо похороненные муравьи тут же снова вернулись в гнездо, но были опять вынесены ревностными поклонниками чистоты. И так происходило до тех пор, пока на теле муравьев оставался «трупный запах».
258
СОДЕРЖАНИЕ
| Предисловие ...................................................................................................................................... |
3 |
| ПОРАЗИТЕЛЬНЫЙ МИР ................................................................................................................. |
5 |
| Первое знакомство ............................................................................................................................ |
7 |
| Путь к социальности ......................................................................................................................... |
16 |
| БИОЛОГИЯ И ОБРАЗ ЖИЗНИ ....................................................................................................... |
29 |
| Общественные осы ........................................................................................................................... |
31 |
| Общественные пчелы ....................................................................................................................... |
47 |
| Пчелы-галикты .................................................................................................................................. |
49 |
| Аллодапиновые пчелы ..................................................................................................................... |
54 |
| Шмели ................................................................................................................................................. |
57 |
| Безжалые пчелы ................................................................................................................................ |
66 |
| Медоносные пчелы ........................................................................................................................... |
75 |
| Муравьи .............................................................................................................................................. |
86 |
| Видовое разнообразие и социальная организация ........................................................................ |
89 |
| Муравьиные жилища ........................................................................................................................ |
102 |
| Питание .............................................................................................................................................. |
116 |
| Термиты .............................................................................................................................................. |
128 |
| Видовое разнообразие и социальная организация ........................................................................ |
130 |
| Жилища термитов .............................................................................................................................. |
135 |
| Питание ............................................................................................................................................... |
140 |
| Основание новых гнезд .................................................................................................................... |
144 |
| ОСНОВНЫЕ ПРИНЦИПЫ СОЦИАЛЬНОЙ ОРГАНИЗАЦИИ ..................................................... |
151 |
| Табель о рангах ................................................................................................................................... |
153 |
| Касты и разделение труда ................................................................................................................. |
167 |
| Жизненный путь ............................................................................................................................... |
187 |
| Государства и федерации .................................................................................................................. |
201 |
| В КРУГУ СЕБЕ ПОДОБНЫХ ....................................................................................................... |
215 |
| Общественный желудок ................................................................................................................ |
217 |
| Внимание к ближнему .................................................................................................................... |
227 |
| Способы общения ............................................................................................................................ |
236 |
| Визитные карточки ......................................................................................................................... |
249 |
| Мобилизация ...................................................................................................................................... |
258 |
| Хлеб насущный .................................................................................................................................. |
277 |
| МЕЖВИДОВЫЕ ОТНОШЕНИЯ ...................................................................................................... |
291 |
| Место под солнцем ........................................................................................................................... |
293 |
| Мой дом — моя крепость ................................................................................................................. |
306 |
| Воришки, нахлебники и паразиты ................................................................................................... |
320 |
| Узурпаторы и рабовладельцы .......................................................................................................... |
328 |
| ИНТЕЛЛЕКТУАЛЬНЫЕ СПОСОБНОСТИ И ГРУППОВОЕ ПОВЕДЕНИЕ ................................ |
349 |
| Границы интеллекта .......................................................................................................................... |
351 |
| Феномен коллектива ......................................................................................................................... |
366 |
| На уровне сверхорганизма ................................................................................................................ |
379 |
| Заключение ........................................................................................................................................ | 396 |
| Рекомендуемая литература ................................................................................................................ |
398 |
| Алфавитно-систематический список русских и латинских названий насекомых ....................... | 399 |