| Главная | Библиотека | Форум | Гостевая книга |
НОВОЕ
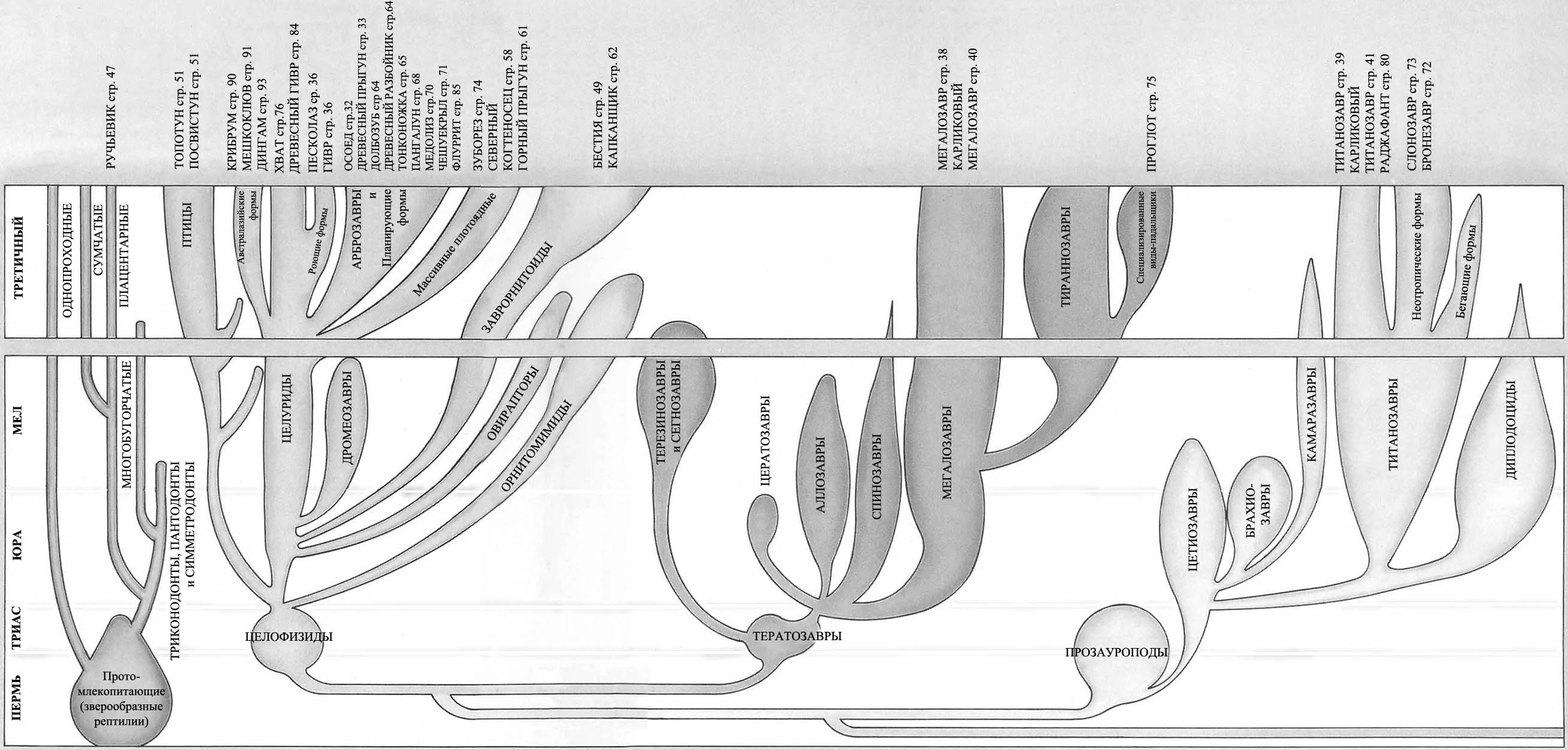
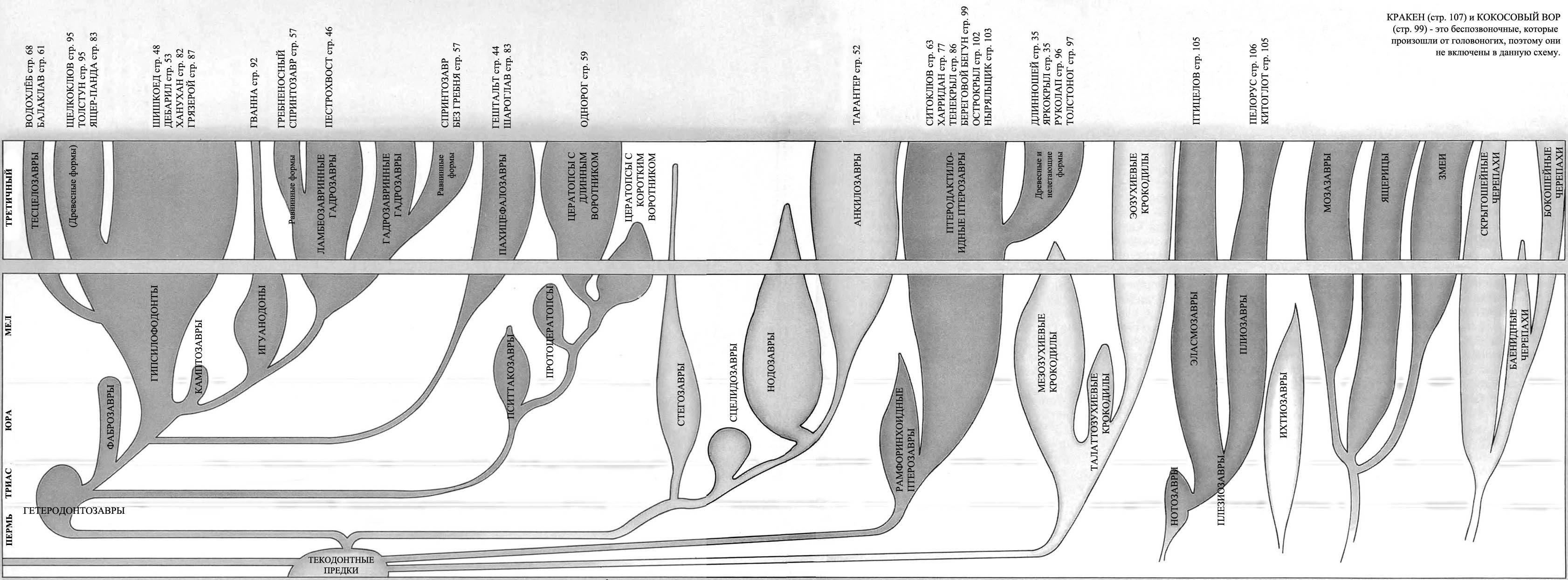
ДРЕВО ЖИЗНИ
Продолжение родословной линии динозавров на протяжении последних 65 миллионов
лет подразумевало, что эти великолепные существа процветали, расселялись
и увеличивали своё разнообразие с той же самой энергией как и на протяжении
предыдущих 150 миллионов лет. Более того, фактически, на протяжении последних
65 миллионов лет они были свидетелями стремительных изменений на поверхности
Земли.
Большинство родословных ветвей динозавров фактически вымерло намного раньше
конца мелового периода, и динозавры, которые живут сегодня, происходят от
наиболее успешных из позднемеловых групп.
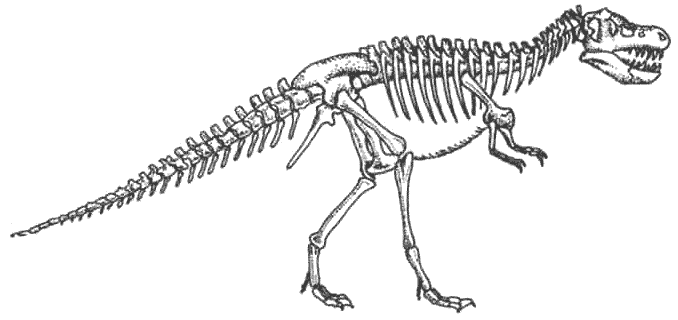
Легко сложенные плотоядные ящеротазовые, или «целурозавры», произошедшие
от целофизид, процветали, будучи очень универсальными и способными к большому
количеству эволюционных изменений.
Плотоядные тяжёлого телосложения, или «карнозавры», произошедшие от тератозавров,
жили не столь хорошо. К концу мелового периода процветали только огромные
тираннозавры. Мегалозавры сохраняют свою численность, но линия карнозавров
не показывает значительного прироста.
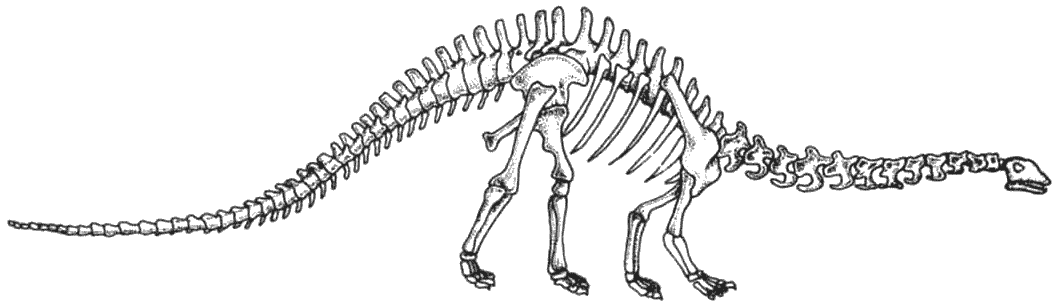
Прозауроподы вымерли в конце триаса, но их потомки – тяжёлые длинношеие растительноядные
ящеротазовые, или «зауроподы», стали обычными в юрский период, но в позднем
мелу существовали только в местах, откуда они не были вытеснены орнитоподами.
Те, что выжили до сегодняшнего дня, представляют собой главным образом потомков
титанозавров и большей частью живут на южных материках.
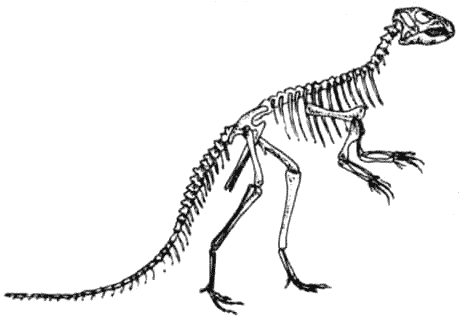
Мелкие двуногие птицетазовые – «орнитоподы» – которые эволюционировали от гетеродонтозавров
(страницы 14 – 15), подобно мелким плотоядным ящеротазовым, адаптировались
и процветали, с лёгкостью приспосабливаясь к значительным изменениям окружающей
среды. Большинство травоядных сегодняшнего дня эволюционировало из их числа.
Цератопсы – рогатые птицетазовые – были очень успешными в конце мелового периода
и всё ещё встречаются в тех или иных формах сегодня.
Стегозавры – птицетазовые, имеющие костные пластины – почти все вымерли к меловому
периоду, за исключением изолированной группы в Индии. Группа не пережила ледниковый
период.
Анкилозавры – покрытые тяжёлой бронёй птицетазовые – были успешной группой
мелового периода и выжили.
Ещё есть летающие птерозавры. Несмотря на конкуренцию с птицами, птерозавры
выжили.
Среди впечатляющих водоплавающих рептилий длинно- и короткошеие плезиозавры,
а также мозазавры выжили, но рыбообразные ихтиозавры вымерли.
А как же млекопитающие? Они начинали в триасовый период как маленькие насекомоядные
существа. К концу мелового периода они всё ещё были маленькими насекомоядными
существами. Они не имели возможностей расселяться и увеличивать разнообразие,
и являются маленькими насекомоядными существами и по сей день*.
* Дугал Диксон сильно обижает млекопитающих своими словами. К концу
мелового периода млекопитающие были очень разнообразными, и он не мог
этого не знать. Среди них существовали однопроходные, сумчатые, насекомоядные,
кондилартры (предки копытных). Возможно, в это же время появились грызуны.
Также существовало несколько архаичных отрядов млекопитающих, в том числе
многобугорчатые, дожившие в нашем мире до эоцена. Самые крупные млекопитающие
мелового периода, возможно, достигали размеров волка. Экология мезозойских
млекопитающих была разнообразнее той, что приписывает им автор книги
– среди них существовали не только насекомоядные формы, но и хищники
и растительноядные. – В. П. |
 |
||||
|
|
|
|
|
 |
|||||||
|
|
|
|
|
|
|
|
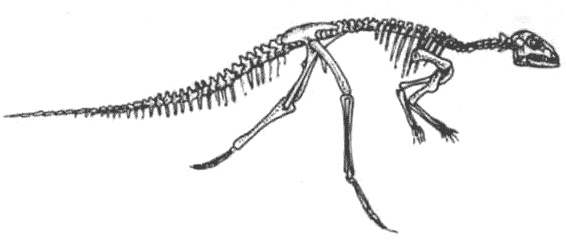
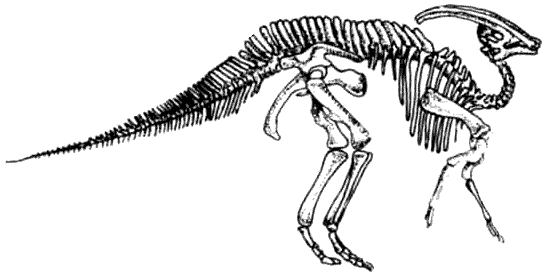
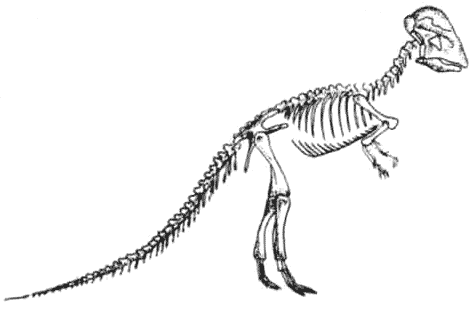
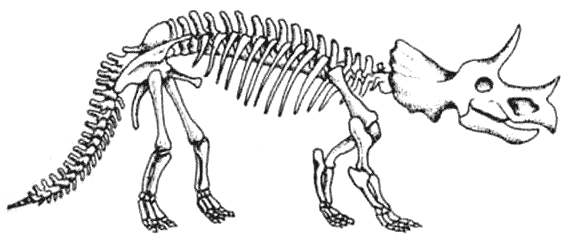
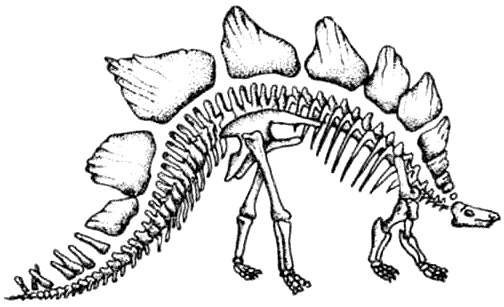
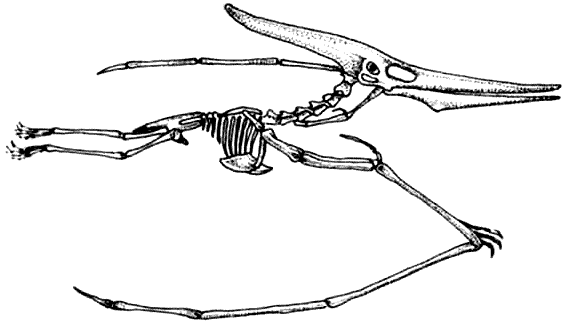
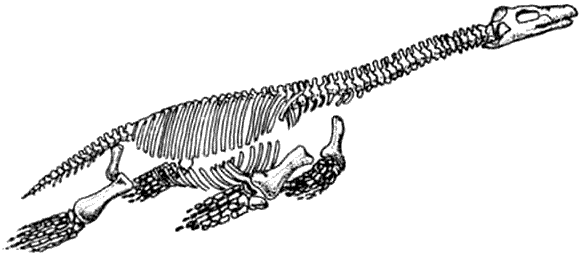
КРАКЕН (страница 107) и КОКОСОВЫЙ ВОР (страница 99) – беспозвоночные, которые происходят от головоногих моллюсков, потому они не включены в эту схему.
ВЕЧНО МЕНЯЮЩИЙСЯ ЛАНДШАФТ
На протяжении основной части Века Рептилий материки раздвигались. В течение
триасового периода все материки Земли были соединены вместе в один огромный
суперконтинент под названием Пангея, состоявший из северной части, Лавразии,
и южной части, Гондваны. В течение юрского периода этот суперконтинент начал
раскалываться, и в меловом периоде разделяющиеся материки отчётливо находились
на пути к их современному положениям.
На протяжении последних 65 миллионов лет, начиная с конца мелового периода,
самое большое движение материков происходило в южном полушарии. В самом конце
мелового периода Австралийский материк всё ещё соединялся с Антарктическим.
Впоследствии он откололся и переместился в северном направлении, пока не
достиг своего нынешнего положения в южном тропическом климатическом поясе.
Континентальный блок, который теперь включает Индию, путешествовал в северном
направлении через Индийский океан, и в конце концов столкнулся с огромным
северным континентом. Поскольку он откололся от своего первоначального положения
как часть восточноафриканского массива суши, он отделил также другие материковые
фрагменты, которые остались после этого как острова, известные как Мадагаскар
и Сейшельские острова, а также множество рассеянных погружённых фрагментов.
Море, называемое Тетис, всё ещё отделяло материки, которые мы теперь называем
Европой и Азией, от Африки, но это море медленно закрывалось и сужалось.
На другой стороне земного шара два американских материка были довольно изолированы,
и связывались только цепочкой островов на протяжении самого раннего третичного
времени, и постоянным сухопутным перешейком, появившимся совсем недавно.
Северная Америка, однако, почти постоянно соединялась с Азией через Берингийский
сухопутный мост – перешеек, который только недавно погрузился в океан.
Движение материков оказало влияние на климат. Когда существовал непрерывный
морской пояс вокруг Земли, состоящий из Тетиса и пролива между Америками,
существовало постоянно текущее на запад экваториальное течение, которое образовывали
господствующие ветры. Оно формировало тёплые и влажные климатические условия
на окраинах большинства материков. Вода, нагретая в этом течении, циркулировала
в океане, доходя вдоль береговых линий до севера северного полушария и далеко
вниз до Антарктиды на юге. Климатические условия были тёплые и ровные, и
влажные леса росли на большинстве материков.
Из-за движения материков море Тетис закрылось. В это же самое время Австралазия
отошла далеко от Антарктиды и открылся циркумполярный морской путь, позволяя
непрерывному течению, направленному на восток, окружить холодный Антарктический
материк. Из-за всего этого пропало экваториальное течение, а вместе с ним
и тёплые климатические условия. Теперь тёплые тропические воды меньше смешивались
с холодными полярными, и климатические условия всё больше и больше различались
на обширных территориях. Более сухие и прохладные условия, ставшие результатом
этих событий, означали, что пышные тропические леса начали уступать место
травянистым равнинам – новой среде обитания.
|
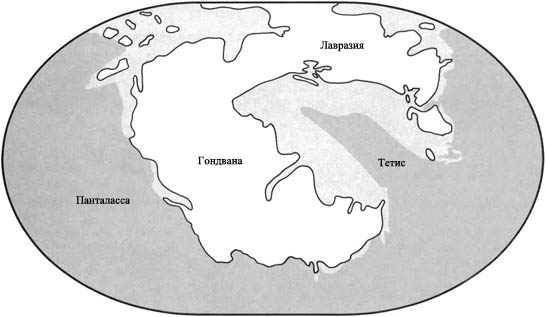
Пангея
Триасовый период был временем, когда суперконтинент Пангея существовал во
свей своей полноте. Он был разделён на две огромных составных части,
каждая часть гораздо крупнее, чем самый большой из сегодняшних материков.
Более северной части, состоящей из современных Северной Америки, Европы
и Азии, дали название Лавразия. Южная часть, состоящая из Южной Америки,
Африки, Индии, Антарктиды и Австралии, называется Гондвана. Морской залив,
называемый Тетис, вдаётся с востока и почти разделяет эти две части.
Остальная территория морей была объединена в мировой океан под названием
Панталасса.
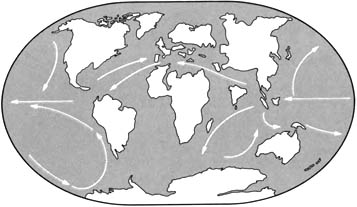 |
Ранний третичный период
Поскольку составные части Пангеи разделились, существовал ярко выраженный
морской путь вокруг Земли, по которому текло на запад экваториальное
течение, изменявшее мировой климат.
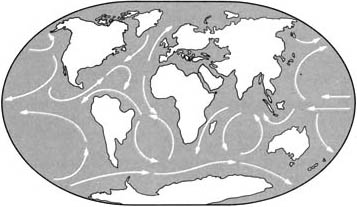 |
Поздний третичный период
Дальнейшие движения материков закрыли экваториальный морской путь и открыли
циркумполярный путь вокруг Антарктиды. Теперь циркуляция воды в океане
не оказывала никакого объединяющего эффекта на общемировой климат.
Климатические условия, постепенно становящиеся холоднее, сформировали ледяной
покров на полюсах Земли. Северный Ледовитый океан был почти полностью окружён
сушей, и постоянный приток рек разбавлял солёную воду. Поскольку перемешивание
с более тёплой водой с юга было небольшим, этот северный океан замёрз насовсем.
В южном полушарии Антарктический материк лежит на полюсе. Этот континент
находился далеко от тёплых океанских течений и был весьма действенно изолирован
от них циркумполярным течением. В результате Антарктида также замёрзла.
Климатические условия продолжали охлаждаться до ледникового периода, приблизительно
1.7 миллионов лет назад. Ледники протянулись на юг от Северного Ледовитого
океана и вниз по склонам гор, а климатические пояса сдвинулись друг на друга
в сторону экватора.
Топография мира в наши дни – прямые последствия этого движения материков.
Когда материк движется, имеется тенденция к образованию горных хребтов вдоль
его переднего края. Самые большие горные хребты Земли – Гималаи вдоль соединения
между Индией и Азией, где эти два материка сблизились и столкнулись 50 миллионов
лет назад, и Скалистые горы и Анды вдоль западных побережий Северной и Южной
Америки, где движение в западном направлении всё ещё не прекратилось. Движение
Африканского материка в сторону Европы создало несколько изогнутых горных
цепей в этой области, вызвав появление гор Атлас, Апеннин, Альп и других
гор, которые окружают Средиземноморье. Цепочка островов Ост-Индии между Азией
и Австралазией может считаться подводными горными хребтами, появление которых
вызвано движением подводных континентальных масс и океанского дна. Многие
другие большие горные хребты мира старые и стёртые – это свидетельства движений
материков в прежние времена. Старые горы Аппалачи по восточному побережью
Северной Америки были когда-то одной непрерывный горной цепью вместе с Шотландскими
горами и Норвежскими горами вдоль края северной Европы, до того, как эти
материки раскололись и разошлись. Урал, старый горный хребет между Европой
и Азией, показывает, где эти два континента столкнулись 300 миллионов лет
назад. Движения, которые разорвали Пангею на части, всё ещё продолжаются.
Когда материк раскалывается, он поднимается горным хребтом, и разламывается
вдоль гребня. Восточно-африканские возвышенности представляют собой такое
поднятие, а трещины присутствуют в виде Восточно-Африканской зоны разломов
(Великой Рифтовой долины), которая тянется вдоль всего материка к открытой
трещине Красного моря.
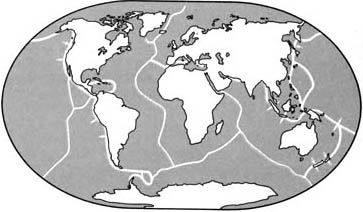 |
Современные движения литосферных плит
Земная кора всё ещё находится в движении, и литосферные плиты всё ещё непрерывно
перемещают материки. Вдоль активных зон, где сходятся литосферные плиты,
могут подниматься горы и вулканы.
Движения материков оказывают большое воздействие на живую природу. Материк может двигаться из одной климатической области в другую, и соответствующим образом фауна и флора должны эволюционировать, чтобы подстраиваться к изменяющимся условиям. В ином случае материк может приблизиться к соседнему массиву суши, и население двух областей смешивается. Все эти виды воздействия находятся в ведении понятия зоогеографии, которая определяет, какие сочетания животных в каких областях земного шара обитают.
РАСПРЕДЕЛЕНИЕ ЖИВОТНЫХ В МИРЕ
Животные не одинаковы по всему миру. Есть множество правил, которые определяют
разновидности животных, которые могут жить в каком-то определённом месте,
но не в другом.
Наиболее явно выраженный фактор, который определяет фауну некоторой местности
– это окружающая среда той местности: климат, рельеф, растения, которые там
растут, другие животные, которые там обитают, фактически всё, что оказывает
влияние на окружающую местность и среду обитания. Мы можем непосредственно
видеть, что животное, которое живет в горной местности, и особенно хорошо
приспособлено для жизни в горной среде обитания, долго не проживёт, если
попадёт в болотистую местность, и наоборот. Выносливость горного животного
при жизни в высокогорье была бы бесполезна в области низинных болот, а обтекаемая
форма болотного животного будет выглядеть весьма неуместной среди скалистых
горных пиков. Но всё же, несмотря на столь глубокое влияние окружающей среды,
горное животное может быть довольно близкородственно болотному животному,
которое живёт на том же самом материке. Они оба могли эволюционировать от
одного и того же предка всего лишь несколько миллионов лет назад. На другом
материке на другом конце мира могут существовать горные животные и животные
из болотистых местностей, которые сходным образом близкородственны друг другу,
но они будут довольно дальними родственниками тем видам, которые есть на
первом материке. Два горных животных могут выглядеть весьма похожими друг
на друга, и два болотных животных будут обладать одним и тем же приспособлением,
но все они будут эволюционировать достаточно независимо.
Объединяя животных мира в группы согласно их эволюционным отношениям, нежели
согласно окружающим условиям, в которых они обитают, можно разделить мир
на удобные для рассмотрения районы, известные как ЗООГЕОГРАФИЧЕСКИЕ ОБЛАСТИ.
Каждая зоогеографическая область будет содержать ряд групп животных, которые
являются специфичными для неё, и которые эволюционировали сравнительно независимо
от совокупности животных другой области. Границы между областями могут быть
явно отмечены, например, океанами, или же могут быть весьма размытыми, когда
несколько типов животных оказываются способны пересечь её и перейти из одной
области в другую. Существование областей является также следствием наличия
естественных преград для миграций, которые появляются из-за физической географии.
Горная цепь или пустыня может отделить одну совокупность животных от другой.
ЭФИОПСКАЯ ОБЛАСТЬ состоит из большей части того, что мы называем Африканским
материком. Северная граница чётко проходит через Сахару и Аравийскую пустыню.
Немногие животные могут пересечь такой барьер, и животные на юге были склонны
развиваться в изоляции от тех, что обитали на севере. Остров Мадагаскар –
часть Эфиопской области, хотя он изолирован от материка и мог бы рассматриваться
как небольшая зоогеографическая область сам по себе.
ПАЛЕАРКТИЧЕСКАЯ ОБЛАСТЬ состоит из материков Европы и Азии к северу от Гималаев.
Она также включает северное побережье Африки. Пустыня Сахара представляет
собой гораздо более серьёзное препятствие для миграций, нежели Средиземное
море.
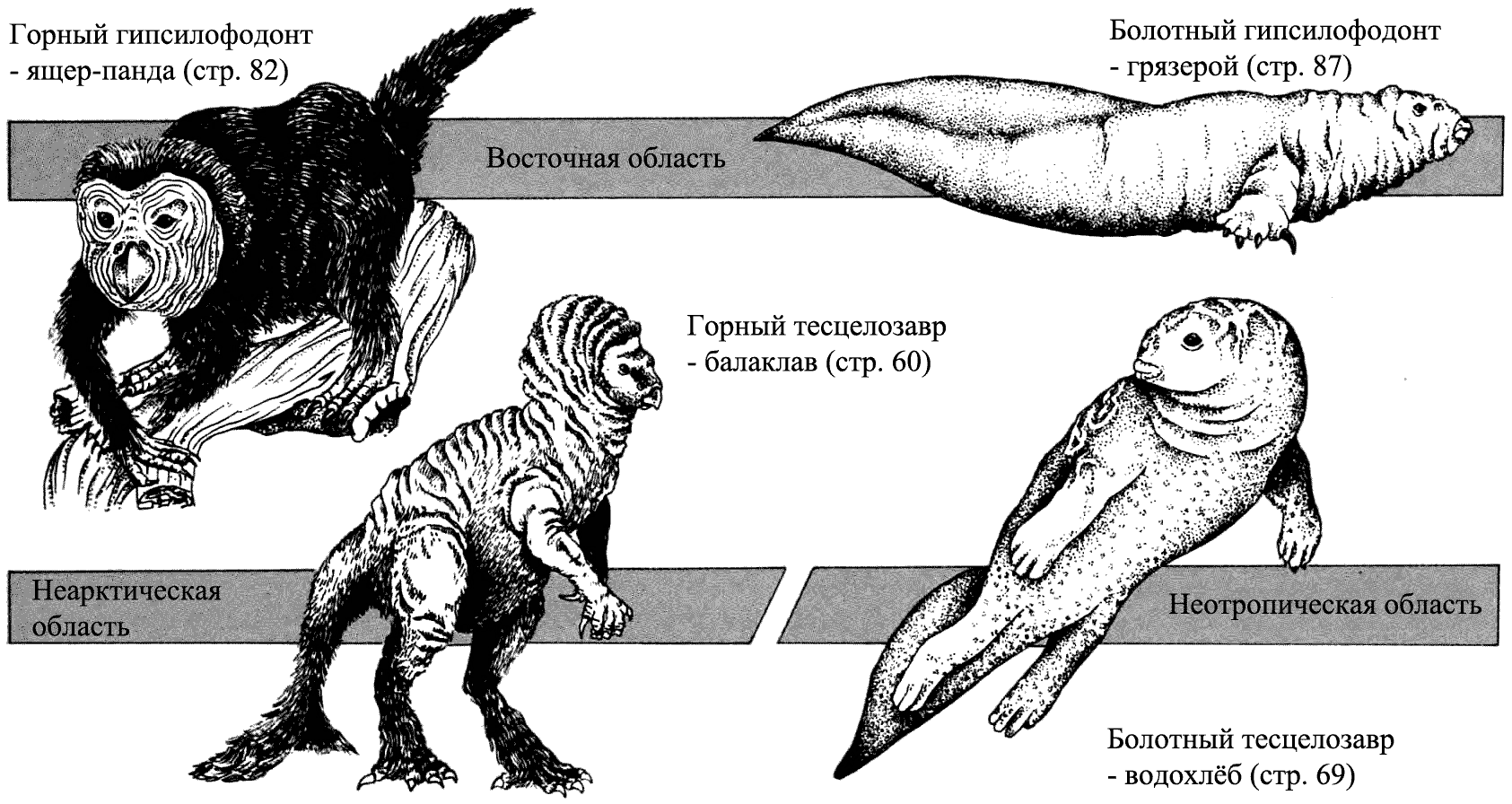
Влияние окружающей среды
В течение мелового периода гипсилофодонты были очень успешной группой. Их
подгруппа – тесцелозавры – эволюционировала в Северной Америке. Балаклав
и водохлёб – два тесцелозавра, потомки западных зоогеографических областей.
Один живёт в горах, другой в болотах, и потому физически они сильно отличаются
друг от друга. В Восточной области ящер-панда и грязерой – потомки основной
линии гипсилофодонтов. Горная форма демонстрирует те же самые приспособления,
что и балаклав, а болотная форма напоминает водохлёба, но они не близкородственны.
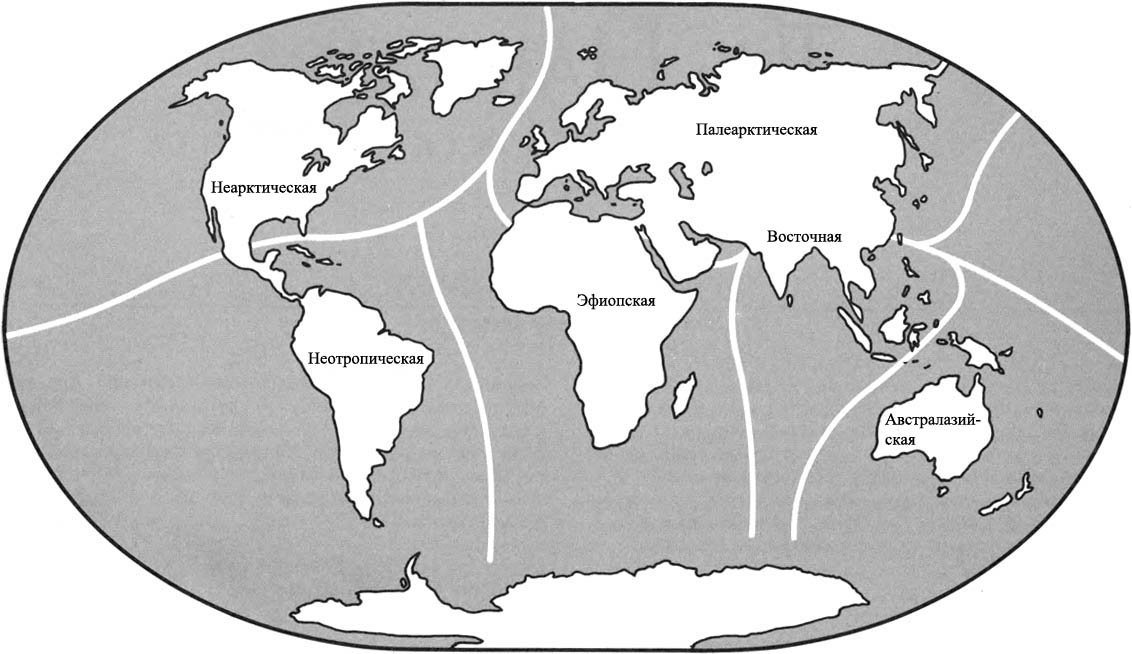 |
Зоогеографические области
Массивы суши современного мира могут быть разделены на шесть фаунистических
зон, или зоогеографических областей, каждая область с совершенно особым
животным населением. Границы каждой области отмечены препятствиями для
миграции и обычно состоят из пустынь, гор или водных пространств. Границы
могут быть отчетливыми или размытыми. Один большой массив суши на земном
шаре – Антарктический материк – настолько беден жизнью, что он не включается
ни в одну из зоогеографических областей.
НЕАРКТИЧЕСКАЯ ОБЛАСТЬ – Североамериканский материк, к северу от Мексиканской
пустыни. Эта пустыня отделяет область от континента на юге. Берингов пролив
отделяет её Палеарктической области на севере и западе, но это весьма непостоянный
барьер, и эти два царства имеют общие особенности.
НЕОТРОПИЧЕСКАЯ ОБЛАСТЬ состоит из Южноамериканского материка и островного
моста Центральной Америки. В течение большей части последних 65 миллионов
лет она была изолированным островом, но совсем недавно она соединилась с
Неарктической областью, и образование сухопутного моста означало, что эти
два царства различаются теперь не так явно, как ранее.
ВОСТОЧНАЯ ОБЛАСТЬ состоит из того, что мы называем Юго-Восточной Азией. Область
Азии к югу и к востоку от Гималаев, включая большинство островов Ост-Индии,
составляет Восточную область. Горы на севере и пустыня на западе обеспечивают
барьеры между этим царством и остальной частью континента.
АВСТРАЛАЗИЙСКАЯ ОБЛАСТЬ, возможно, наиболее чётко изолирована среди всех
областей. Она была островным континентом с тех пор, как откололась от Антарктического
материка, и растительная и животная жизнь развивались там по их собственному
пути.
К этому списку из шести зоогеографических областей мы добавили седьмую, это
ОКЕАНЫ. В отличие от континентов, океаны не имеют границ, и между одним океаном
и другим не существует никаких физических барьеров. Фауна, существующая в
океанах, имеет тенденцию к широкому распространению и основывается скорее
на температурах и условиях водной среды – иными словами, на окружающей среде
– нежели на эволюционном развитии в каких-либо областях. На материках совокупности
животных, водящихся в каждой области, отражают историю той области. Они зависят
от того, когда и как фауна мигрировала в ту отдельно взятую область, и от
последующего развития условий окружающей среды, таких, как географические
и климатические факторы, которые влияют на животных, живущих в той местности.
В мире, в котором выжили динозавры и другие мезозойские существа, фауна всё
равно подчинена ограничениям современной зоогеографии. Соответствующим образом
разделена и эта книга. Без вымирания рептилий не было никакого развития фауны
млекопитающих, и никакой эволюции людей, а потому и никакого обучения географов.
Следовательно, местам в мире не было дано названий, под которыми они известны
сегодня. Поэтому, ради целей этой книги, мы обошлись без обычных географических
названий, которые основаны на культурном и политическом разграничении и отражают
исследования и историю человечества, и повсюду в тексте мы будем обращаться
к местностям только в контексте их зоогеографических областей*.
* Можно добавить, что границы и названия зоогеографических областей могут
варьировать у разных авторов и научных школ. Так, Мадагаскар может выделяться
из Эфиопской области в самостоятельную Мадагаскарскую, а Палеарктическая
и Неарктическая области входят в единую Голарктическую на правах отделов.
Восточная область в России чаще всего носит название Индо-Малайской, но при
переводе я отдал предпочтение названию, менее связанному с географическими
названиями, поскольку, в соответствии с идеей этой книги, автор предпочитает
не использовать названия, данные людьми, кроме самых необходимых. Строго
говоря, при условиях, выдвинутых Диксоном, не должно быть ни Палеарктики,
ни Неарктики – ведь «арктос» означает «медведь», а в его мире динозавров
никто не назвал созвездие в честь никогда не появлявшегося зверя, и, соответственно,
области в честь этого созвездия!
Флористическое районирование суши гораздо более дробное, нежели фаунистическое
– в частности, на территории Африки выделяется до шести флористических областей,
и имеется Голантарктическое флористическое царство. – В. П.
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ
ВЕЛИКОЕ ВЫМИРАНИЕ 6
ЧТО ТАКОЕ ДИНОЗАВР? 10
НОВОЕ ДРЕВО ЖИЗНИ 12
ПАЛЕОГЕОГРАФИЯ 16
ЗООГЕОГРАФИЯ 18
СРЕДЫ ОБИТАНИЯ 20
НОВЫЕ ДИНОЗАВРЫ 29
ЭФИОПСКАЯ ОБЛАСТЬ 30
ПАЛЕАРКТИЧЕСКАЯ ОБЛАСТЬ 42
НЕАРКТИЧЕСКАЯ ОБЛАСТЬ 54
НЕОТРОПИЧЕСКАЯ ОБЛАСТЬ 66
ВОСТОЧНАЯ ОБЛАСТЬ 78
АВСТРАЛАЗИЙСКАЯ ОБЛАСТЬ 88
ОКЕАНЫ 100
ЗАКЛЮЧЕНИЕ 108
ПОСЛЕСЛОВИЕ 109
СЛОВАРЬ 113
СОВЕТУЕМ ПРОЧИТАТЬ 115
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ 116
БЛАГОДАРНОСТИ 120