ИСТОРИЯ
ЖИЗНИ
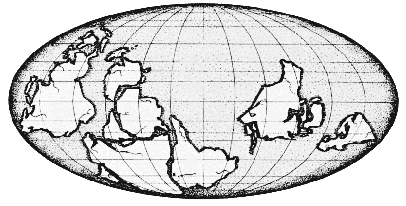
На карте показано расположение материков в начале кембрия, точки во времени, начиная с которой более отчётливо отслеживается история жизни. В докембрийское время многие животные были мягкотелыми и сохранялись очень редко.
Земля существует около 5000 миллионов лет и населена организмами
того или иного рода около 3500 – 4000 миллионов лет из этого отрезка времени.
Тем не менее, точная летопись окаменелостей земной жизни может быть отслежена
примерно до 620 миллионов лет назад – до времени, когда впервые появились
твёрдые скелеты. В это время жизнь существовала только в море, а суша была
безжизненна.
Распределение суши и моря было не таким, как в наши дни. Расположение материков
и океанов постоянно меняется из-за процесса, который называется тектоника
плит. Земная кора состоит из совокупности плит, напоминая части футбольного
мяча. Эти плиты постоянно нарастают вдоль одного края, где составляющее их
вещество поднимается из недр Земли, и разрушаются на другом краю, где плита
поддвигается под соседнюю и расплавляется.
Поднятие дна происходит вдоль серединно-океанических хребтов, а разрушение
идёт вдоль глубоководных желобов. Материал, вовлечённый в эти процессы, представляет
собой океаническую кору, богатую окисью кремния и магнезитом. Континенты
состоят из иной разновидности коры, богатой окисью кремния и алюминием, которая,
будучи легче, находится на поверхности, и таким образом континенты движутся
в разных направлениях по земному шару с помощью тектонической активности.
Этот процесс происходил на протяжении всего геологического времени, и будет
продолжаться, пока существует мир. Важность тектоники плит для истории жизни
на Земле не ограничивается только географией. Тектоника плит частично оказывает
влияние на характер глобального климата, который меняется в течение сравнительно
коротких в геологическом смысле отрезков времени, и, несомненно, вносит свой
вклад в относительно внезапные изменения, которые происходят с доминирующими
формами жизни на Земле. Взаиморасположение континентов на ключевых стадиях
развития этих животных было в какое-то время важным для их распространения
по Земле, и стало причиной явных различий между формами жизни, населяющими
разные массивы суши.
Солнце и солнечная система сформировались из бесформенного облака межзвёздного газа, медленно вращающегося в космосе со скоростью около одного оборота за десять миллионов лет. Вращаясь, оно начало сокращаться под влиянием своей собственной силы тяготения, и вследствие этого, начало вращаться быстрее. Силы, участвующие в этом процессе, сплющили газовое облако в диск, в котором вещество начало собираться в центре, формируя Солнце. Ближе к краям диска образовались завихрения, которые начали собирать вещество, формируя зачатки того, что позже станет планетами. Частицы пыли, состоящие главным образом из вкраплений железа и частиц соединений кремния, начали объединяться. Частицы слипались в сгустки и собирались вместе в завихрениях под действием гравитации. Железо, будучи тяжелее, погружалось в середину, а окись кремния оставалась на поверхности, давая протопланетам железное ядро, окружённое каменной мантией. Таким путём формировались внутренние планеты – Меркурий, Венера, Земля и Марс. Остальные планеты формировались из лёгких материалов, таких, как углекислый газ и аммиак, которые конденсировались из газа, когда температура продолжала падать. В это время сжатие материи в раннем Солнце запустило процесс слияния ядер, и Солнце начало излучать энергию – этот процесс продолжается последние 5000 миллионов лет, и будет продолжаться ещё около 5000 миллионов лет.
В эпоху, предшествующую времени точных научных исследований, человек верил, что Земля в том виде, в котором он её знал, и все живые существа, которых он знал, были результатом единственного сверхъестественного акта творения, который произошёл в определённое время в сравнительно недавнем прошлом. Ископаемые морские животные, находимые на суше далеко от моря, позже признанные свидетельством значительных изменений в очертаниях суши и моря, были объявлены результатом карающего потопа. |
Возможно, первая атмосфера Земли содержала много водорода, метана и аммиака,
подобно составу атмосферы внешних планет Солнечной системы. Со временем к
этим газам добавились водяные пары и углекислый газ, выделяющиеся из новообразующихся
гор. Вода вначале оставалась в виде пара, пока жар атмосферы не позволял
ей конденсироваться. С другой стороны, столь же вероятно, что первичная атмосфера
из водорода, метана и аммиака была большей частью «сдута» излучением солнца
вскоре после её образования, и позже первая стабильная земная атмосфера была
сформирована преимущественно из углекислого газа и водяного пара, выделившихся
из недр через фумаролы и вулканы. В любом случае вода, конденсировавшаяся
и выпадавшая в виде дождя, когда Земля остыла, несомненно содержала молекулы
аммиака, метана и водорода, растворённые в ней. Когда этот раствор подвергался
высокоэнергетическому воздействию вроде ударов молний или ультрафиолетового
излучения солнца, могли происходить химические реакции, которые способствовали
синтезу сложных органических молекул вроде аминокислот – материал, из которого
состоят живые существа.
С другой стороны, возможно совершенно иное объяснение происхождения сложных
органических молекул. Простые органические соединения вроде формальдегида
имеются в межзвёздной пыли – частицах углерода, образующихся при взрывах
звёзд. Молекулы этих органических веществ могли аккумулироваться на частицах
и постепенно объединяться в длинные химические цепочки сложных органических
молекул, которые представляют собой первый шаг в химии живого. Газ, выделившийся
из звёзд, может содержать кислород, углерод и азот. Если газ содержит больше
кислорода, чем углерода и азота, могут формироваться органические молекулы
типа полисахаридов (простые сахара). Если преобладающим элементом является
азот, более вероятно образование нуклеиновых кислот и хлорофилла – воспринимающего
энергию вещества зелёных растений. Межзвёздная пыль может под воздействием
сил гравитации собираться в сгустки, и при стечении обстоятельств попадать
на околосолнечную орбиту в виде комет. Если такая комета столкнулась с Землёй
в ранние эпохи её формирования, что более чем вероятно, межзвёздные органические
молекулы могли достигнуть поверхности нашей планеты.
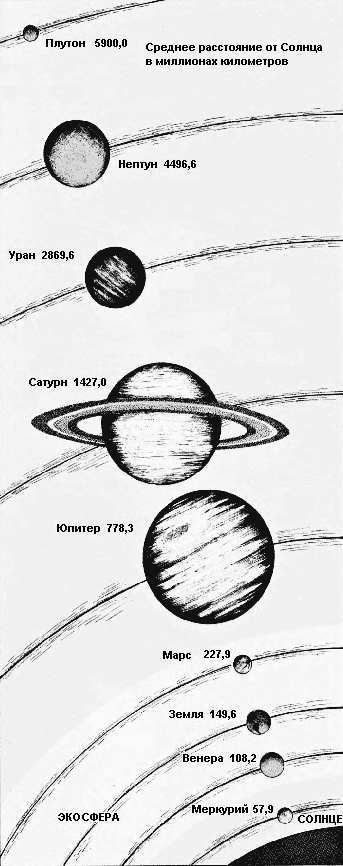
Подходящие условия в атмосфере и на поверхности делают возможным существование жизни на планете, которая лежит в пределах пояса вокруг Солнца, который называется экосфера. Этот пояс простирается от расстояния немного ближе орбиты Венеры до расстояния немногим дальше орбиты Марса. Меркурий, где максимальная температура на поверхности 370°С, слишком нагрет, чтобы поддерживать жизнь, а внешние планеты, становясь всё холоднее вплоть до Нептуна и Плутона, где максимальная температура ниже - 200°С, слишком холодны.
|
Как бы то ни было, очевидно, что горячие моря на жаркой поверхности Земли
4500 миллионов лет назад содержали сложные органические молекулы, необходимые
для построения и развития живых существ.
Первым объектом на Земле, который мог наверняка называться «живым», была
молекула с уникальной способностью к самовоспроизводству. Чтобы это сделать,
она должна быть способна разрушать сложные молекулы, такие, как полисахариды,
и использовать их составные части для построения собственной зеркальной копии.
Любая особенность исходной молекулы, помогавшая ей в решении этой задачи,
повышала бы её шансы на выживание, и такая особенность могла сохраняться
в процессе самовоспроизводства. Любая особенность, которая замедляла этот
процесс, вела бы к вымиранию молекулы. Эволюция началась.
Такая деятельность продолжалась до тех пор, пока исходные полисахариды, находившиеся
в «первичном бульоне», не были использованы полностью. Прото-организмы могли
дальше остаться без пищи, если бы они не выработали способность синтезировать
свою собственную органику из неорганических веществ, используя солнечную
энергию. Этот процесс, известный как фотосинтез, стал возможным благодаря
наличию молекул хлорофилла.
Со временем в объект репликации стало включаться больше, чем одна сложная
молекула, и так появилась компактная органическая структура, известная как
клетка. Некоторые из наиболее примитивных клеток не имели центрального ядра,
места, где находится аппарат репродукции клетки, а вместо этого данная функция
была рассеяна по цитоплазме. Также существовали клетки с ядрами, у которых
было гораздо большее будущее; в ходе эволюции мелкие клетки объединились
в более крупные образования, оставаясь в них, чтобы выполнять какие-то жизненно
важные межклеточные функции. Постепенно в сложных образованиях, состоящих
более чем из одной клетки, каждая клетка стала играть свою особую роль в
поддержании жизни всего объединения. Эволюционировал организм.
Эволюция первых многоклеточных организмов могла идти одним из двух возможных
путей. Либо свободноживущие клетки разных типов объединились в одну структуру,
либо клетки прекратили полностью разделяться во время деления и сохранялись
вместе как единое целое. Безотносительно способа их образования, эти многоклеточные
организмы должны были быть более успешными, нежели сумма их частей; в противном
случае они бы не выжили.
Клетки многоклеточных организмов не одинаковы, и имеют довольно различающиеся
функции в зависимости от тканей и органов, которые они образуют. У высших
форм жизни некоторые из них являются строительными элементами, подобно клеткам
костной ткани, другие, такие, как клетки крови, обеспечивают защиту от болезней
и перенос питательных веществ, тогда как иные, нервные клетки, образуют системы
органов чувств и связи в организме. Дифференцировка клеток в большинстве
случаев происходит на стадии зародыша. Вначале клетки зародыша одинаковы.
Первоначальная оплодотворённая яйцеклетка делится на две дочерних клетки,
которые делятся на четыре, и так до тех пор, пока не образуется несколько
сотен одинаковых клеток. Как бы то ни было, на определённой стадии развития
зародыша эта стадия прекращается, и образуются специфические клетки, предназначенные
для того, чтобы выполнять в организме определённые роли. Неясно, как происходит
эта дифференцировка клеток. Все ядра клеток содержат одинаковую генетическую
информацию, но только часть её используется при образовании новой клетки.
Какие-то факторы внутри клетки, вероятнее всего, внутри клеточного ядра,
должны определять, какая часть генетического кода используется для построения
новой клетки таким образом, чтобы она могла выполнять функцию, присущую ей.
В мире беспозвоночных существует две формы симметрии: радиальная (A), при которой животные симметричны относительно оси, проходящей вдоль тела от вершины до основания, и двусторонней, при которой животные симметричны относительно плоскости, проходящей вдоль их тел.* * На рисунке показаны брюхоногие моллюски, у которых двусторонняя
симметрия нарушена из-за превращения раковины в турбоспиральную (башенковидную).
Но это преобразование вторично – у примитивных моллюсков явная двусторонняя
симметрия. – В. П. |
В ранних океанах процветали одноклеточные и многоклеточные существа, растения
и животные. Растения были способны поглощать солнечную энергию и фотосинтезировать
пищу из неорганического материала. Животные, не способные самостоятельно
производить пищу с помощью солнечного света, получали энергию, поедая растения.
Это различие в способах питания является основным различием между растениями
и животными, и отражается на строении и физиологии этих двух типов организмов.
Растениям, нуждающимся только в солнечном свете и неорганических веществах,
нет необходимости двигаться, оказавшись в благоприятных условиях, и потому
их клетки твёрдые и имеют жёсткие стенки. У них есть плоские поверхности,
поглощающие энергию (листья), которые направлены в сторону солнца, и удерживающие
структуры (корни), через которые они поглощают питательные вещества, и которые
предохраняют растение от того, чтобы быть сдутым или смытым водой. С другой
стороны, животные в большинстве случаев должны передвигаться от растения
к растению, и потому они выработали в процессе эволюции более гибкие клеточные
стенки и систему мускулов, чтобы сделать передвижение возможным. Они развили
органы чувств и нервную систему, с помощью которых они оценивают своё окружение
и передают сигналы мускулам.
Геометрия самого животного связана с его способностью двигаться. Те, кто
не является почти бесформенными сидячими комками, фильтрующими пищу из проходящего
течения воды, имеют либо радиальную, либо двустороннюю симметрию.
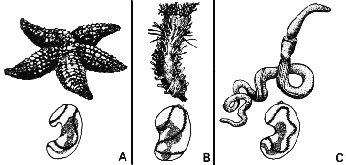
Баланоглосс (C) Balanoglossus spp. – это полухордовое, промежуточная стадия между беспозвоночными и хордовыми – группой, которая включает позвоночных. Сходство между личинками баланоглосса и личинками морской звезды (A) и голотурии (B), которые обе являются иглокожими, может означать наличие у хордовых беспозвоночных предков. |
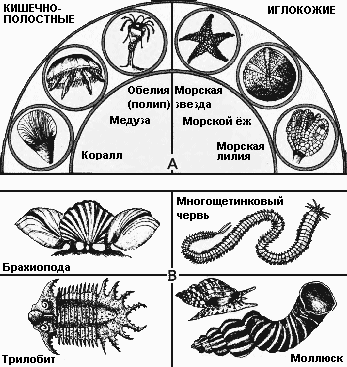
В начале кембрийского периода впервые в большом количестве появились животные с твёрдыми покровами. Поскольку в норме сохраняются в ископаемом виде только панцири животных, дальнейшая история жизни становится хорошо известной только с этого времени. В кембрии эволюционировали все крупные группы (типы) животных, и радиально-, и двусторонне-симметричные. Животные с радиальной симметрией включают кишечнополостных (медузы и кораллы) и иглокожих (морские звёзды и морские ежи).* Формы, имеющие двустороннюю симметрию, распадаются на четыре главных группы: плеченогие – почти вымершая группа раковинных животных; моллюски – двустворчатые, морские улитки и похожие на наутилусов головоногие; членистоногие – представленные преимущественно трилобитами; а также несколько классов червей и червеобразных животных.
| * Радиальная симметрия иглокожих вторична и развилась в связи с переходом к пассивному образу жизни. Древнейшие кембрийские иглокожие были двусторонне-симметричными, а некоторые формы вообще не имели осей или плоскостей симметрии. По происхождению иглокожие не связаны с кишечнополостными и близки к хордовым, в том числе позвоночным, о чём упомянуто в тексте. – В. П. |
От одной из групп этих червеобразных животных, хордовых, в силуре произошли
первые животные с позвоночником – класс примитивных бесчелюстных рыб и предки
всех позвоночных. Также в это время растения впервые вышли на сушу. Из прибрежных
мелководий вышла группа растений, которые могли выживать, не будучи полностью
погруженными в воду. В процессе эволюции у них развились жёсткие стебли,
чтобы обеспечивать им опору, и внутренняя проводящая система, чтобы доставлять
воду и растворённые минеральные вещества из земли, и переносить произведённую
пищу от листьев.
Как побочный эффект фотосинтеза в атмосферу выделялся свободный кислород; содержание
кислорода увеличивалось, тогда как содержание углекислого газа уменьшалось,
делая состав воздуха более подходящим для жизни животных. Членистоногие были
первыми животными, которые получили преимущество от улучшения условий в атмосфере,
и скорпионы с многоножками обитали среди ранних растений.
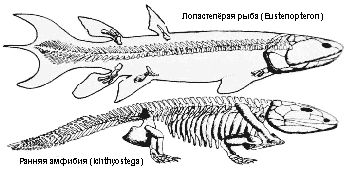 Сходство
между лопастепёрыми рыбами, такими, как Eustenopteron, и ранними
амфибиями, как Ichthyostega, демонстрирует явное свидетельство
происхождения амфибий. У Ichthyostega рыбий позвоночный столб из
однородных позвонков был замещён более тяжёлым и прочным образованием
и полностью развитой грудной клеткой, позволяющими поддерживать
животное на суше. Её конечности, хотя и длиннее, чем рыбьи плавники,
похожи на них по форме. Сходство
между лопастепёрыми рыбами, такими, как Eustenopteron, и ранними
амфибиями, как Ichthyostega, демонстрирует явное свидетельство
происхождения амфибий. У Ichthyostega рыбий позвоночный столб из
однородных позвонков был замещён более тяжёлым и прочным образованием
и полностью развитой грудной клеткой, позволяющими поддерживать
животное на суше. Её конечности, хотя и длиннее, чем рыбьи плавники,
похожи на них по форме. |
Членистоногие были первыми животными, которые получили преимущество от улучшения
условий в атмосфере, и скорпионы с многоножками обитали среди ранних растений.
Следующий девонский период известен как Век Рыб. Первыми эволюционировавшими
от примитивных бесчелюстных форм были плакодермы (пластинокожие) вроде динихтиса
(Dinichthys) – панцирные рыбы, которые имели челюсти, эволюционировавшие
из костей жаберных дуг. До конца девона они были большей частью замещены
хрящевыми рыбами типа кладоселахии (Cladoselache), предшественниками акул
и скатов. Костные рыбы, наиболее разнообразные и широко распространённые,
существовали параллельно этим хрящевым видам. Они делились на две главных
группы – лучепёрые рыбы, которые стали наиболее успешными, и лопастепёрые
рыбы, такие, как Eustenopteron. Последняя группа наиболее важна из этих двух
с эволюционной точки зрения. Обитание в мелких пресных водоёмах, которые
периодически пересыхали, дало им эволюционный стимул выживать вне воды.*
Когда водоёмы исчезали, Eustenopteron полз по суше к соседнему водоёму с
помощью пары мускулистых плавников, развившихся из органов-стабилизаторов.
В это время он был способен дышать воздухом с помощью примитивных лёгких,
развившихся из выростов глотки. Жизнь позвоночных на суше началась, даже
хотя она была лишь временной мерой, позволяющей продолжить существование
в воде. К концу девона появились амфибии, способные проводить большую часть
взрослой жизни на суше. Одна из ранних, ихтиостега, демонстрирует типичное
строение пятипалой конечности, поддерживаемой сильными поясами конечностей,
известными у наземных животных. Но всё равно она сохраняла рыбьи особенности
в форме хвоста и черепа.**
| * В настоящее время существует иная точка зрения на происхождение земноводных
– они могли быть потомками рыб, освоивших густо поросшие растениями болота,
а лапы развились не для движения на суше, а для ползания среди зарослей.
– В. П. ** На момент написания этой книги конечность ихтиостеги не была известна и реконструировалась по более поздним находкам. У ранних позвоночных конечность несла не пять, а больше пальцев – у Tulerpeton их было шесть, а у Acanthostega восемь. – В. П. |
Следующий каменноугольный период был временем великих угольных лесов. Он
также был Веком Земноводных; обильно заросшие болота, характерные в этот
период для низменностей, были идеальными для их развития, и вследствие этого
появилось большое количество их новых видов. Некоторые, вроде долихозомы
(Dolichosoma), были мелкими и похожими на угря, для других, таких, как эогиринус
(Eogyrinus), предполагается внешность и образ жизни, как у аллигатора. Ещё
одни, наподобие диплокаулуса (Diplocaulus), стали широкими и уплощенными,
и жили исключительно в иле. Черепа этих животных были более развитыми, чем
рыбообразный череп ихтиостеги. Носовые отверстия были хорошо оформившимися,
что говорит о том, что они принадлежали к числу приспособленных к дыханию
воздухом животных. Эти животные дали начало более поздним продвинутым амфибиям,
а также рептилиям.
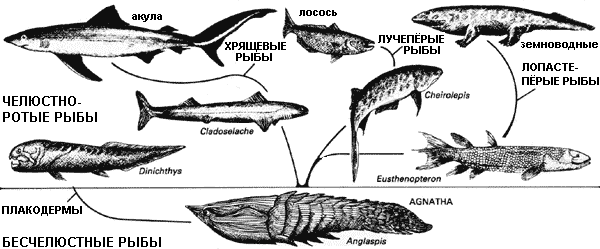
Ранние рыбы были бесчелюстными (агнатами), их рот представлял собой не более, чем отверстие пищеварительного тракта. Челюстноротые рыбы впервые появились в девоне. Наиболее примитивные, плакодермы, были очень разнообразной группой панцирных видов с челюстями и зубами, сформировавшимися из костных пластинок головы. Хрящевые рыбы, предки акул и скатов, также появились в это время. Костные рыбы, наиболее успешная группа, также произошедшие от бесчелюстных, могут быть разделены на два класса: лопастепёрые рыбы, имеющие мясистые плавники, и лучепёрые рыбы, которые имеют плавники, состоящие из кожи, поддерживаемой роговыми лучами. Большинство видов рыб, живших в Эпоху Человека, принадлежало к классу лучепёрых. Лопастепёрые рыбы были представлены только четырьмя родами.*
* Диксон считает лопастепёрых и лучепёрых рыб отдельными классами. Существует
много взглядов на систематику рыб, и не все авторы поддерживают точку зрения
Диксона.
Современные лопастепёрые рыбы – это три рода двоякодышащих рыб, рогозуб (Neoceratodus),
лепидосирен (Lepidosiren) и протоптерус (Protopterus), а также кистепёрая
рыба латимерия (Latimeria). Ранее был известен один современный вид с Коморских
островов, но недавно был открыт ещё один вид из вод острова Сулавеси. – В.
П.
Рептилии были первыми полностью наземными позвоночными на Земле. Амфибии,
от которых они произошли, были достаточно хорошо приспособлены к жизни на
суше, но всегда должны были возвращаться в воду для размножения, и на ранних
стадиях развития всегда неминуемо должны были вести полностью водный образ
жизни головастика. Это означает, в сущности, что колонизация земноводными
суши ограничивается болотистыми местностями около побережий, озёр и речных
берегов.
Развитие рептилий, превзошедшее этот уровень - это развитие яйца с твёрдой
оболочкой, которое с помощью непроницаемых плёнок позволило зародышу животного
развиваться в своём собственном бронированном водоёме индивидуального пользования
вдали от воды. В дополнение к этому у рептилий есть плотная шкура, которая
препятствует высыханию в горазд большей степени, нежели шкура земноводных.
Ранние рептилии, известные как «стволовая группа рептилий», произошли от земноводных в каменноугольный период и развились во множество форм, которые заполнили все главные среды обитания – воздух, сушу и воду. Ихтиозавры, плезиозавры и мозозавры были водными, птерозавры были обитателями воздуха, а динозавры и зверообразные рептилии были наземными. Динозавры («ужасные ящеры») классифицируются на две группы согласно строению их таза. Как ни странно, птицы произошли от группы ящеротазовых, а не от птицетазовых, как можно было бы ожидать. Как часто бывает в природе, когда недостаток изощрённости заключает в себе способность к адаптации, тип строения крокодила, одна из рано эволюционировавших форм тела рептилий, в конечном итоге доказала свою успешность. |
Хотя первые рептилии появились среди угольных лесов каменноугольного периода, рептилии реально вступили в свои права не ранее пермского и триасового периодов. Поскольку в это время большие участки суши становились гораздо более сухими, возможность жить вдалеке от воды превратилась в важное преимущество; это прекрасные условия, способствующие расселению и многообразию рептилий.
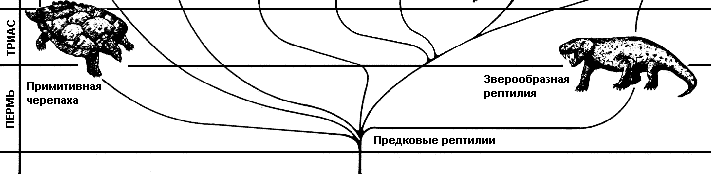
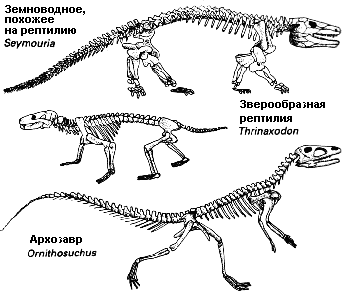
От рептилиеобразных земноводных типа сеймурии (Seymouria) произошли две группы наземных рептилий – архозавры и зверообразные пресмыкающиеся. Ранние архозавры вроде орнитозуха (Ornithosuchus) были двуногими, но многие поздние представители группы приспособились к четвероногому передвижению. Зверообразные рептилии, например, тринаксодон (Trinaxodon), хотя появились рано в эволюционной истории рептилий, выжили, чтобы стать предками млекопитающих. |
Первой действительно успешной группой рептилий были зверообразные. Их зубы
представляли собой значительный шаг вперёд по сравнению с простыми зубами-капканами
для рыбы у амфибий. У них были длинные смертоносные зубы в передней части
рта, и более короткие режущие зубы в задней; это приспособление к хищническому
образу жизни было прообразом специализированной зубной системы млекопитающих.
Конечности сдвинулись в характерное для млекопитающих положение под туловище
таким образом, что оно опиралось на вершины ног, нежели болталось между ними,
что характерно для амфибий и ранних рептилий. В конце триасового периода*
зверообразные рептилии вымерли, оставив в качестве своих потомков настоящих
млекопитающих, которые полностью реализовали свой потенциал в течение последующих
100 миллионов лет. Тем временем ниша доминирующих рептилий была захвачена
группой, известной как архозавры, к которой позднее принадлежали динозавры.
Архозавры («правящие рептилии») впервые появились в пермский и триасовый
периоды как полуводные существа, по форме слегка похожие на крокодилов, с
мощными задними лапами и сильными хвостами – признаками, которые сохранялись
на протяжении истории всей группы. Когда в пермский период эти полуводные
рептилии вновь приспособились к жизни на суше, они обнаружили, что их длинные
хвосты дают достаточное равновесие, чтобы позволить им ходить на задних ногах
– эволюционировало основное двуногое положение динозавров.
| * Остатки зверообразных рептилий известны из более поздних отложений – тритилодонтов (Tritylodontidae) находят в юрских отложениях до конца средней юры, а остатки Chronoperates известны из раннего палеоцена (кайнозой), но неясно, был ли он высокоразвитой зверообразной рептилией, или же примитивным млекопитающим. – В. П. |
К началу юрского периода динозавры стали доминирующей формой позвоночных
и дали значительное многообразие форм, которые распространились по всем пригодным
для обитания территориям земной суши. Появились крупные травоядные, такие,
как диплодок (Diplodocus), легко сложенные быстроногие хищные формы типа
дейнониха (Deinonychus) и более медлительные питающиеся мясом падальщики
вроде аллозавра (Allosaurus). Ошибочно полагать, что все динозавры были массивными
зверюгами. Компсогнатус (Compsognathus), питавшийся главным образом мелких
рептилий и яйца, был не крупнее домашней курицы. Легко сложенные травоядные
динозавры, типичным представителем которых был камптозавр (Camptosaurus),
были особенно важной группой и были предками нескольких крупных групп, особенно
примечательных панцирных разновидностей конца мелового периода. Защитные
приспособления динозавров приобретали разнообразные формы и размеры в зависимости
от своих функций: вертикальные пластины, как у стегозавра (Stegosaurus),
плоские костные наросты, как у анкилозавра (Ankylosaurus) и головные щиты,
как у трицератопса (Triceratops).
Ни одна из ранее упомянутых групп архозавров не подтвердила в итоге свою
успешность. В случае с любой группой животных существует тенденция, при которой
наиболее примитивные и неспециализированные представители наиболее выносливы,
и в случае архозавров самой долгоживущей группой оказались ранние крокодилы,
которые пережили множество эффектно выглядевших динозавров более, чем на
сто миллионов лет, выжив с некоторыми изменениями и после Эпохи Человека.
В своё время архозавры властвовали в небе. Самыми первыми летунами были птерозавры,
существа, скользившие по воздуху на кожистых крыльях, растянутых между удлинённым
четвёртым пальцем их передних конечностей и их задними ногами и хвостом.*
Среди них были мелкие животные типа птеродактиля (Pterodactylus) и рамфоринха
(Rhamphorhynchus), которые могли питаться насекомыми, более крупные формы
вроде птеранодона (Pteranodon) и никтозавра (Nyctosaurus), которые были рыбоядными,
и чудовищные существа наподобие кетцалькоатля (Quetzalcoatlus), которые почти
наверняка были падальщиками.
| * Диксон опирается в описании на ранние реконструкции птерозавров, когда их считали неким подобием летучих мышей и реконструировали соответствующим образом. Более поздние авторы считают, что задние ноги птерозавров могли быть свободны от летательной перепонки. – В. П. |
В юрский период другая ниша летающих существ была занята потомками одной
особой группы мелких плотоядных динозавров, похожих на компсогнатуса (Compsognathus),
у которых были перья вместо чешуи, покрывавшие, по крайней мере, часть их
тел (скорее всего, их конечности и хвосты). Они были предками настоящих птиц,
которые стали хозяевами небес в более позднюю кайнозойскую эру.*
Во время периода эволюции динозавров другие группы рептилий развивались в
формы, способные использовать обильные пищевые ресурсы, существовавшие в
море. Чтобы вернуться в царство своих далёких предков, они вынуждены были
повторно развить многие физические особенности, утраченные в процессе приспособления
к жизни на суше.
| * Происхождение птиц от динозавров – не столь однозначный вывод. Находка
триасового протоависа (Protoavis) заставляет предположить, что эволюция
птиц началась раньше и может быть не связана с динозаврами, а открытие
группы «не настоящих» птиц энанциорнисов (Enantiornitiformes) указывает
на то, что превращение рептилий в птиц шло несколькими родословными линиями
параллельно. – В. П. ** Хвост ихтиозавров по строению не похож на хвосты рыб. У акул и осетров позвоночник продолжается в верхнюю лопасть хвоста, а у ихтиозавра – в нижнюю. – В. П. |
Группа, которая в Эпоху Человека включает змей и ящериц, в то время дала
начало тилозавру (Tylosaurus), морскому хищнику с массивными челюстями, который
двигался в толще воды с помощью волнообразных изгибов своего длинного тела
и уплощенного хвоста, и использовал веслообразные конечности для поворотов.
Плезиозавр (Plesiosaurus), медленно плавающее существо с длинной змеевидной
шеей, которую он использовал, чтобы внезапно атаковать и ловить рыбу, свою
главную пищу, не имеет выживших потомков. Венцом творения в эволюции водных
рептилий, возможно, был ихтиозавр (Ichthyosaurus), рептилия, которая выглядела
скорее как рыба. У него было тело обтекаемой формы, плавники и рыбий хвост.**
Несмотря на эти замечательные приспособления крупные рептилии полностью вымирают
к концу мелового периода. Причина этого до сих пор неясна, но их вымирание
совпадает с резкими изменениями фауны морских беспозвоночных и с климатическими
изменениями, результатом которых была смена типов растительности на большей
части Земли.
Хотя млекопитающие эволюционировали от зверообразных рептилий в триасовый
период, они оставались маленькой незначительной частью фауны позвоночных
следующие 100 миллионов лет, пока рептилии правили бал на суше, в море и
воздухе.
Все характерные черты строения млекопитающих – упрощение нижней челюсти,
эволюция уха из челюстных костей, специализированное строение зубов и положение
конечностей под телом, а не по сторонам от него – выработались уже у зверообразных
пресмыкающихся. Черта строения, которая особенно чётко отделяет зверообразных
рептилий от млекопитающих – это сочленение нижней челюсти. Челюсть рептилии
– это сложно устроенное образование, состоящее из нескольких костей. У млекопитающих,
однако, челюсть состоит из одной кости. Дополнительные кости, имеющиеся у
рептилий, объединены в ухе млекопитающего. Развитие в этом направлении происходило
независимо по меньшей мере, четыре раза в конце эры господства зверообразных
рептилий.
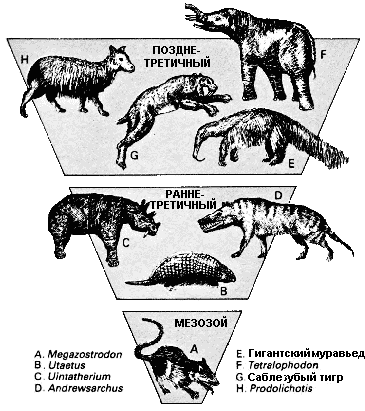
Все млекопитающие произошли от мелких насекомоядных зверьков, таких, как мегазостродон (Megazostrodon). В конце мезозоя результатом крупных изменений в окружающей среде было вымирание динозавров, и млекопитающие эволюционировали, чтобы занять их место как доминирующая группа позвоночных. Раннетретичные млекопитающие были главным образом лесными жителями. Хищники этого периода, как эндрюсарх (Andrewsarchus), были примитивными и имели зубы, похожие на зубы рептилий. Когда в конце позднего третичного периода условия стали более сухими, леса отступили, и появились млекопитающие, подобные тем, которые известны в Эпоху Человека. |
Некоторые физиологические особенности, характерные для млекопитающих, также возникли у зверообразных рептилий. Нёбо, которое даёт млекопитающим возможность дышать и есть одновременно, важное для постоянного снабжения кислородом, необходимого для поддержания теплокровного состояния, впервые возникло у зверообразных рептилий. Ямки на черепе и челюстных костях, указывающие на наличие вибриссов, позволяют заключить, что некоторые зверообразные рептилии были хотя бы частично покрыты волосами и опять является свидетельством теплокровности. Развитие их зубов указывает, что молодняк был беззубым, и, следовательно, матери должны выкармливать его молоком.
| * Новейшие находки мезозойских млекопитающих не подтверждают этого.
Среди них известны более крупные формы – размером с лисицу. Кроме того,
среди млекопитающих мезозоя были специализированные формы – водные, например,
касторокауда (Castorocauda), и планирующие – волантотерий (Volantotherium).
Млекопитающие развивались в пределах своего размерного класса, конкурируя
с молодью динозавров. В конце мелового периода уже существовали даже
архаичные копытные. – В. П. ** Отряд многобугорчатые (Multituberculata) вымер только в эоцене – это самый долгоживущий отряд млекопитающих. – В. П. |
На протяжении Века Рептилий разнообразие млекопитающих ограничивалось мелкими,
похожими на мышь существами, питавшимися насекомыми, семенами, а также, возможно,
яйцами рептилий.* Хотя в юрский период существовало несколько различных отрядов,
различающихся по строению зубов, немногие из них пережили динозавров.** Наиболее
успешными мастерами выживания были плацентарные – те млекопитающие, которые
снабжали питанием развивающегося детёныша внутри своего тела, пока он не
достигал более высокой стадии развития. Плацентарные впервые появились в
меловой период параллельно боковой ветви развития – сумчатым, млекопитающим,
которые выкармливают своё развивающееся потомство в наружной сумке. Сумчатые
и плацентарные вместе образуют почти всю мировую фауну млекопитающих в Эпоху
Человека, хотя есть группа примитивных яйцекладущих млекопитающих, однопроходные
(клоачные), которая представлена лишь горсткой видов, включая утконоса.
Расцвет Века Млекопитающих начался в раннем третичном периоде, с уходом крупных
рептилий. В течение первых десяти миллионов лет или около того все ниши,
ранее занятые рептилиями, был заполнены млекопитающими, и появились все отряды
млекопитающих, известные в Эпоху Человека.
На суше травоядные были представлены
копытными, например, лошадями и свиньями, у которых были зубы для срезания
и пережёвывания растений, крупными слонами
с остроконечными бивнями и мелкими грызунами и зайцеобразными типа крыс и
кроликов, с грызущими передними и перемалывающими задними зубами. На этих
травоядных охотились свирепые представители семейства хищников. Насекомых
и других беспозвоночных поедали примитивные насекомоядные, землеройки, с
крепкими зубами, способными пробить твёрдые наружные покровы жуков и многоножек,
и неполнозубые, муравьеды и броненосцы, вовсе без зубов.* Приматы, низшие
и человекообразные обезьяны, эволюционировали в лесах и питались широким
спектром кормов, что нашло отражение в универсальном характере строения их
зубов. Все эти млекопитающие – плацентарные, но на некоторых изолированных
материках, в Австралии и Южной Америке, вместо них многие ниши были заняты
сумчатыми.
| * Диксон не совсем корректен в этом обобщении. У броненосцев есть зубы, и у некоторых вымерших форм (например, Peltephilus) они были очень крупными. Специализированный насекомоядный броненосец Stegotherium имел рудиментарные зубы. – В. П. |
В воздухе ниша рептилий птерозавров была занята летучими мышами, крылья
которых развились из вытянутых передних конечностей и пальцев. Поскольку
они летают преимущественно вечером и ночью, они не конкурируют непосредственно
с птицами, которые гораздо лучше приспособились к такому образу жизни.
Водные ниши плезиозавров, плиозавров и ихтиозавров были заняты, соответственно,
тюленями, китами и дельфинами, у которых в процессе эволюции развились плавники
и веслообразные конечности.
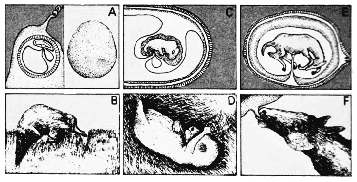 Млекопитающие
делятся на три больших группы – однопроходные, сумчатые и плацентарные.
Однопроходные (A), представленные утконосами, откладывают яйца.
У самок нет оформленных молочных желез, и, когда вылупляется молодняк,
он слизывает молоко матери с поверхности её живота (B). Сумчатые
(C) рождают недоразвитый молодняк, который в большинстве случаев
находится и питается в брюшной сумке (D). Плацентарные (E), как
говорит их название, имеют плаценту, через которую зародыш питается
до тех пор, пока не разовьётся достаточно хорошо. После того, как
детёныши рождаются, мать выкармливает их с помощью развитых молочных
желез (F). Млекопитающие
делятся на три больших группы – однопроходные, сумчатые и плацентарные.
Однопроходные (A), представленные утконосами, откладывают яйца.
У самок нет оформленных молочных желез, и, когда вылупляется молодняк,
он слизывает молоко матери с поверхности её живота (B). Сумчатые
(C) рождают недоразвитый молодняк, который в большинстве случаев
находится и питается в брюшной сумке (D). Плацентарные (E), как
говорит их название, имеют плаценту, через которую зародыш питается
до тех пор, пока не разовьётся достаточно хорошо. После того, как
детёныши рождаются, мать выкармливает их с помощью развитых молочных
желез (F). |
Первая половина третичного периода характеризуется увеличением числа различных
видов млекопитающих. Это выглядело так, словно природа испытывала все разновидности
новых форм, чтобы увидеть, какая из них лучше подходит к свободной экологической
нише. Позже разнообразие млекопитающих свелось к гораздо меньшему числу лучше
приспособленных видов. Это аналогично ситуации, последовавшей за исчезновением
человечества и вымираниями, которые вызвал непосредственно он. Вновь, почти
немедленно, эволюционировало большое количество видов, а позже разнообразие
сократилось до значительно меньшего числа успешных форм.
Раннетретичные наземные млекопитающие были лесными обитателями. Затем примерно
в середине периода глобальный климат изменился, и в умеренных и тропических
широтах раскинулись травянистые равнины, предоставлявшие больше открытых
биотопов. Злаки, составляющие основу растительности этих мест, представляют
собой богатый и неистощимый источник корма, но прежде, чем его можно будет
начать использовать, должны эволюционировать складчатые перетирающие зубы,
чтобы справиться с высоким содержанием в нём кремнезёма. Открытое пространство
делает маскировку сложнее и вынуждает травоядных млекопитающих, населяющих
его, спасаться от опасности бегством, и, чтобы оставаться с добычей, хищники
также должны быть быстрее.
Приматы, которые эволюционировали на деревьях в лесу, также вторглись на
травянистые равнины. Одна их особая группа выработала выпрямленное положение
тела – естественное продолжение их способа передвижения по деревьям – и это
позволило им смотреть поверх высокой травы и предупреждать о приближающихся хищниках. Другим
наследством их древесной жизни была замечательная координация руки и глаза.
Эта способность позволила им использовать палки и камни как инструменты,
чтобы более эффективно использовать доступную пищу. Особенностью приматов,
которая особенно помогла им в сборе пищи, был высокий уровень их социальной
организации. Охотясь группами, они могли окружать животное, которое в ином
случае могло быть слишком осторожным и быстроногим, чтобы его можно было
поймать. Повышение уровня интеллекта, которое позволило им координировать
эти особенности поведения и регулировать сложную социальную структуру, в
будущем стало путём эволюции человека.
 |
 |
 |
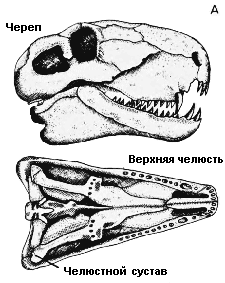
В ходе эволюции
от рептилии к млекопитающему челюсть стала более усовершенствованной;
челюстной сустав сдвинулся дальше вперёд, чтобы обеспечить более
точное жевание; зубы стали специализированными и приобрели различную
форму, чтобы служить разным целям, таким, как откусывание, жевание
и нанесение ран. Челюстной сустав рептилии стал соединяться со
средним ухом, и у млекопитающих развилось нёбо, так что они могут
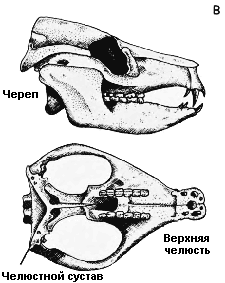
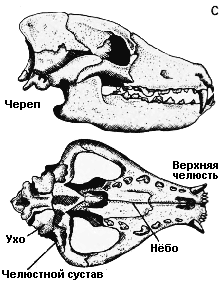
дышать и есть одновременно. A. Ранняя зверообразная рептилия диметродон (Dimetrodon); B. Поздняя зверообразная рептилия олигокифус (Oligokyphus); C. Млекопитающее собака (Canis) |
||
Первые люди были собирателями растений и охотниками на животных, и мало отличались по образу жизни от остальных травоядных и хищных животных вокруг них. У них были достаточный интеллект, чтобы делать орудия труда и оружие, и общественная организация, которая делала более эффективными охоту и сбор пищи. Ни одна из этих вещей тем не менее, не наносила какого-либо серьёзного ущерба окружающей среде.*
| * Это не так. В настоящее время учёные склоняются к выводу, что вымирание множества представителей мегафауны на разных материках связано с охотничьей деятельностью человека. Вымирание крупных животных в Новом Свете и Австралии совпадает во времени с заселением этих частей света человеком. – В. П. |
Первое великое изменение в их образе жизни произошло, когда вместо охоты и сбора животных и растений они собрали их в одном месте и начали ухаживать за ними. Это устранило элемент опасности, связанный с охотой, и уменьшило вероятность голода, потому что больше не было вероятности возвращения из похода за пищей с пустыми руками. Это было началом сельского хозяйства.
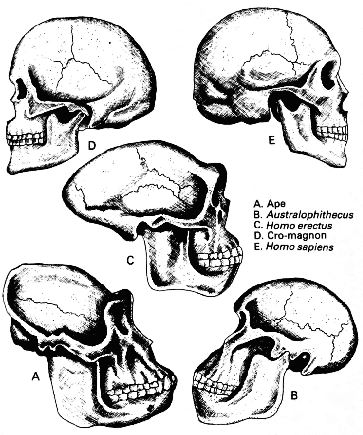
Эволюция человека может быть прослежена от обезьяноподобного предка из числа гоминид, австралопитека (Australopithecus) и ранних представителей нашего собственного рода вроде человека прямоходящего (Homo erectus). Кроманьонец, ранняя форма вида Homo sapiens как такового, появился в Европе к концу плейстоценовых оледенений. Череп человека развивался из массивного образования, заключающего в себе маленький мозг, в череп из более лёгких костей, формирующих крупную мозговую полость. Большой мозг человека наделил его силой сознательного мышления и отделил его от остального животного царства. |
Первое время сельскохозяйственные территории были маленькими и относительно
незначительными. Тем не менее, улучшения в образе жизни первого человека
были столь разительными, что его популяции стремительно увеличивались, и
всё больше и больше суши приходилось очищать от естественной растительности,
чтобы освободить место для посадок и пасущихся животных.
По мере того, как человеческие изобретательность и способность к изготовлению
орудий труда росли, он изобрёл промышленное производство, с помощью которого
можно было производить инструменты с большей скоростью и меньшими трудностями,
чем ранее. Это неизбежно требовало энергии, и ради обеспечения топливом леса
были сведены на дрова, а горы были раскопаны, чтобы добраться до угля,. За
несколько тысяч лет земные ландшафты были изменены до неузнаваемости.
Человеческое знание росло, особенно значительно в области медицинских наук.
Несчастные случаи и болезни, которые помогали сдерживать рост естественных
популяций, были побеждены, или их влияние было уменьшено усилиями человека.
Генетические дефекты, которые в дикой природе могли оказаться смертельными
и были бы отсеяны естественным отбором, множились, поскольку их носители
имели возможность жить и размножаться. Мировое народонаселение росло в геометрической
прогрессии, и вряд ли хоть какая-то область земного шара осталась не тронутой
человеком.
Конечный эффект этого состоял в том, что, в то время, как другие животные
изменяются и адаптируются в ходе медленного процесса эволюции, чтобы соответствовать
особенностям своих мест обитания, человек способен менять своё окружение,
чтобы оно соответствовало его текущим потребностям, пожиная сиюминутные плоды
преимуществ этого процесса. Существуя вне эволюции, каждая стадия в его стремительном
культурном развитии передавалась следующему поколению не через гены, а посредством
обучения. Хотя человек избежал неприятных последствий естественного отбора,
он также оказался лишён его долговременных благоприятных результатов, и в
скором времени эволюция применительно к нему остановилась. Результатом этого
был мир, отягощённый популяцией существ, неспособных выжить без их собственного
разумного вмешательства, мир, отданный на удовлетворение нужд человека, мир,
отравленный его отходами.
В конце концов, Земля не могла далее оставаться источником сырья, необходимого
для человеческого сельского хозяйства, промышленности и медицины, и, когда
нехватка сырья вызвала упадок одной области деятельности за другой, их комплексное
и взаимосвязанное социальное и технологическое единство рухнуло. Человечество,
более не способное к адаптации, бесконтрольно устремилось к собственному
неизбежному вымиранию.
Когда исчезла доминирующая жизненная форма, мир животных погрузился в период
эволюционного хаоса, который длился десятки тысяч лет. Тем не менее, вымирание
человека дало толчок к образованию множества новых видов животных, и его
исчезновение имело первоочередную важность для формирования облика мира,
который появился 50 миллионов лет спустя.
ПРЕДИСЛОВИЕ Десмонда Морриса 9
Генетика клетки : Естественный отбор : Поведение животных : Форма и Развитие : Цепи питания
ИСТОРИЯ ЖИЗНИ 22
Происхождение Жизни : Ранние формы жизни : Эра рептилий : Эра млекопитающих : Эпоха человека
Мир после Человека
ЛЕСА И СТЕПИ УМЕРЕННОГО ПОЯСА 36
Кролопы : Хищники : Жители подлеска : Древесные жители : Ночные животные : Заболоченные земли
Травоядные млекопитающие : Охотники и их добыча : Жизнь на деревьях
Мигранты : Мичинг и его враги : Северный Ледовитый Океан : Южный Океан : Горы
Обитатели песков : Крупные животные пустынь : Североамериканские пустыни
Поедающие траву : Гиганты равнин : Мясоеды
Полог леса : Живущие на деревьях : Подлесок : Жизнь в воде
Австралийские леса : Подлесок австралийских лесов
ОСТРОВА И ОСТРОВНЫЕ КОНТИНЕНТЫ 100
Южноамериканские Леса : Южноамериканские пампасы : Остров Лемурия
Острова Батавия : Острова Пакаус
Участь жизни
Словарь : Древо Жизни : Указатель: Благодарности