Филип
Дж. Сентер
Филип
Дж. Сентер | Главная |
Из: Skeptical Inquirer том
41,
номер 4, июль/август
2017 г.
Перевод: П. Волков, 2019 г.
Ведущие геологи и биологи признают весьма многочисленные физические свидетельства того, что возраст Земли составляет миллиарды лет, что все организмы – эволюционные потомки общего предка, и что нептичьи динозавры вымерли шестьдесят пять миллионов лет назад (см., например, Gradstein et al. 2004; Prothero 2007). В противоположность им, авторы из числа младоземельных креационистов (МЗК) долгое время утверждали, что сообщение в Книге Бытия о сотворении мира и библейской хронологии верно в буквальном смысле, и относят сотворение Земли и всех типов живых организмов ко времени около 6000 лет назад. Из этого положения делается заключение о том, что динозавры и люди были сотворены в один и тот же день, и потому взаимодействовали друг с другом. Заявление о том, что легенды о драконах основаны на таких встречах, долгое время было оплотом литературы МЗК, и в 1977 году биохимик и МЗК Дуэйн Гиш поднял эту концепцию на новую ступень в своей детской книге «Динозавры, эти ужасные ящерицы» (Dinosaurs, Those Terrible Lizards), утверждая, что динозавры были огнедышащими. Другие авторы из числа МЗК последовали его примеру (см. ссылки ниже), и теперь динозавры дышат огнём в учебниках биологии для седьмого класса издательства BJU Press (Batdorf and Porch 2013; Lacy 2013).
В поддержку идеи о том, что реальное животное способно образовывать огонь, Гиш (1977) упомянул защитный механизм жуков-бомбардиров (Brachinus spp.), которые распыляют в морду потенциальным хищникам смесь перекиси водорода и гидрохинона. Химические катализаторы заставляют смесь разогреваться до обжигающих 100º C (Aneshansley et al. 1969). Последующие авторы МЗК последовали примеру Гиша, и добавили воображаемые детали вроде искр, взрывов или пламени (Phillips 1994; Hamp 2000; Isaacs 2010; Paul 2010). В действительности же жуки просто выбрызгивают горячую жидкость – которая ошпаривает, но не создаёт пламени, и потому совершенно не является биологическим прецедентом в пользу природной способности зажигать огонь.
Некоторые авторы из числа МЗК упоминали в качестве биологического прецедента для зажигания огня биолюминесцентных животных и электрических угрей (Morris 1984; Petersen 1986; Morris 1988; Niermann 1994; Morris 1999; DeYoung 2000; Petersen 2002). Тем не менее, процессы, которые вызывают биолюминесценцию (Haddock et al. 2010) и биологическую генерацию электричества (Pough et al. 2013), химически не родственны горению, и выделяют мало тепла, или его вовсе не выделяется. Поэтому данные процессы не относятся к процессу воспламенения, и совершенно не представляют собой биологического прецедента для него.
Сторонники гипотезы огнедышащих динозавров предложили целый ряд потенциальных механизмов воспламенения, рассмотренных ниже, и каждый из них – отдельная самостоятельная гипотеза. Ниже я воспользуюсь научными данными для оценки каждой из гипотез. Каждая такая гипотеза косвенно предсказывает, что механизм не только возможен физически, но и не причиняет животному серьёзных повреждений. Любая из гипотез оказывается фальсифицированной, если какое-то из этих предсказаний не выполняется.
Выдох самовоспламеняющегося газа
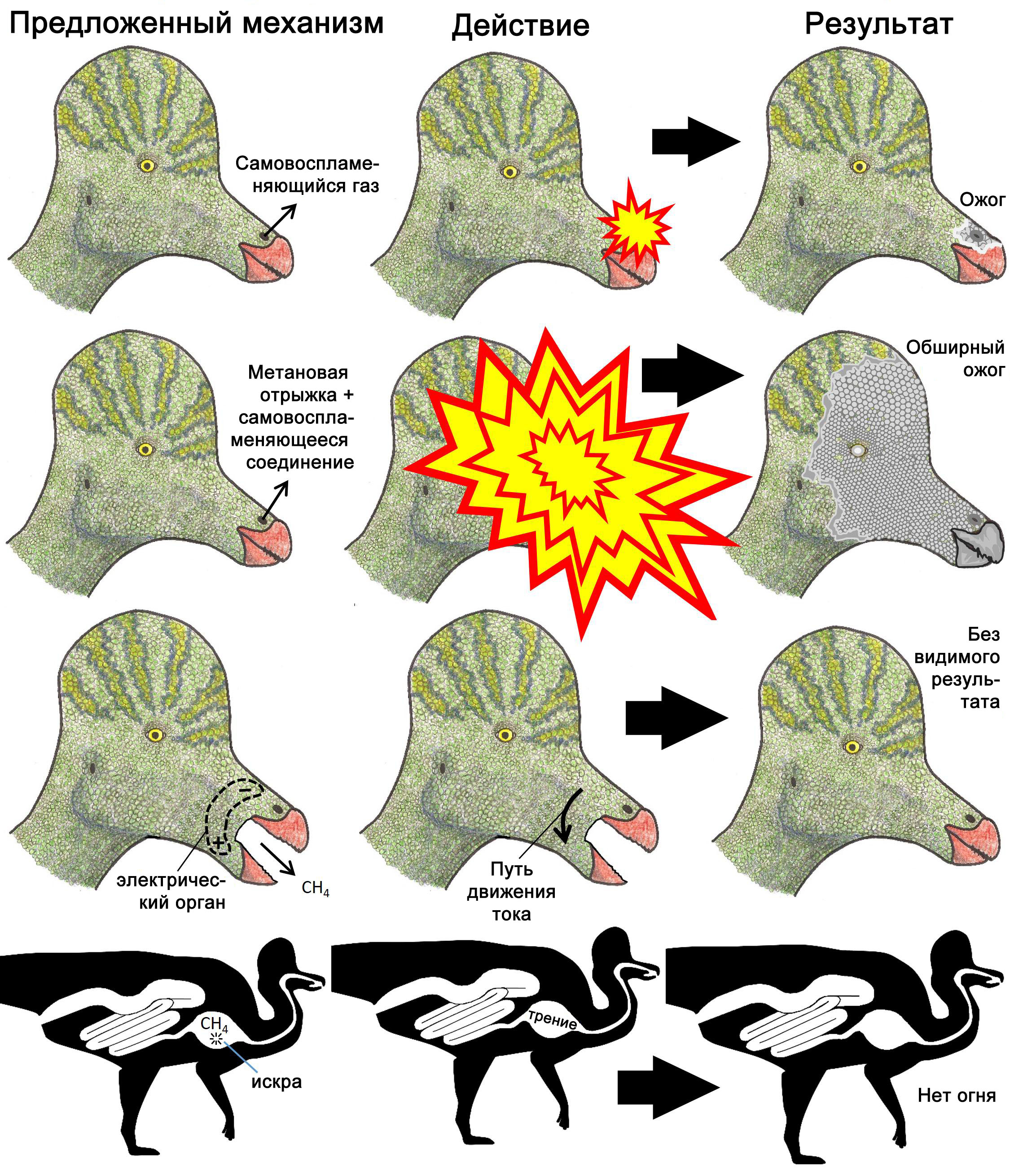
Генри Моррис (1984) и Джеймс Гилмер (2011) предположили, что огнемечущая рептилия могла выдыхать газы, которые воспламеняются после контакта с кислородом. Вещество, которое загорается после контакта с воздухом, называют самовоспламеняющимся (пирофорным). Выпущенный в воздух из ёмкости, самовоспламеняющийся газ взрывается, проделав путь всего лишь в несколько сантиметров или миллиметров от отверстия. Чем шире отверстие, тем ближе к нему происходит взрыв. Например, самовоспламеняющийся газ силан (SiH4) даёт взрыв на расстоянии 5-80 мм от края трубки диаметром 4,32 мм и на расстоянии 5-30 мм от края трубки диаметром 3,5 мм (Tsai и др. 2010). Если оперировать этими данными, то самовоспламеняющийся газ, выпущенный из ноздри или горла крупного динозавра – отверстия диаметром больше, чем несколько миллиметров – взорвался бы немедленно (рисунок 1), обжигая морду или горло животного. Серьёзный вред, который это нанесло бы животному, фальсифицирует данную гипотезу разжигания огня.
Гипотеза могла бы быть состоятельной, если бы ноздри были защищены огнестойкой тканью, но у животных не образуется никаких тканей такого рода. Многие животные образуют сильно кератинизированные производные эпидермиса (волосы, перья, роговые чехлы и т.д.), которые защищают от трения, но горят, когда подвергаются действию огня; то же самое происходит с кутикулами из коллагена или хитина. Если морды динозавров образовывали защитный негорючий минеральный покров (например, из карбоната или фосфата кальция), этот покров должен был сохраняться в ископаемом состоянии, как обычно происходит с твёрдыми частями тел животных. Такой покров полностью отсутствует на морде любой из окаменелостей динозавров.
Воспламенение отрыгиваемого метана
Травоядные млекопитающие ежедневно испускают большой объём эруктации (отрыжки) метана; это происходит в среднем двадцать шесть раз в час у крупного рогатого скота и сорок два раза в час у овец (Colvin et al. 1958; Ulyatt et al. 1999; Koch et al. 2009). Джон Моррис (Morris 1999) предположил, что травоядные динозавры вели себя аналогичным образом, и выделяли из железы самовоспламеняющийся материал для поджигания отрыгиваемого метана.
Однако, будучи высвобожденным из ёмкости (вроде ротовой или носовой полости динозавра), газ немедленно распространяется во всех направлениях; метан быстро окружил бы голову динозавра. Самовоспламеняющийся газ не только вызвал бы ожог носовой или ротовой полости в момент выделения, но также поджёг бы облако насыщенного метаном воздуха вокруг головы животного, обжигая образующимся в результате огненным шаром поверхность головы (рисунок 1). Серьёзный вред, который это могло бы причинить, фальсифицирует данную гипотезу разжигания огня.
Случаи с людьми подтверждают, что возгорание воспламеняющейся отрыжки вызывает ожоги лица. Эруктация у людей обычно включает испускание проглоченного воздуха, который лишён метана и не горюч. Однако в случаях пилоростеноза у взрослых людей, когда затруднён проход в двенадцатиперстную кишку, из-за брожения пищи в желудке в животе могут накапливаться горючие газы. Когда пациент отрыгивает эти газы во время курения, зажжённая сигарета поджигает газы, и образующийся в результате огненный шар вызывает ожоги лица (Galley 1954; MacDonald 1994).
Воспламенение метана электрическим органом
У некоторых видов рыб есть электрические органы, состоящие
из видоизменённых клеток мускулатуры, называемых электроцитами, которые скомпонованы
в ряды таким образом, что их напряжение суммируется (Gallant et al. 2014; Sillar
et al. 2016). Многие электрические виды рыб генерируют слабые, безопасные импульсы,
которые используются для ориентации, обнаружения других рыб и передачи социальных
сигналов (Lissmann 1958; Sillar et al. 2016). В противоположность им, электрические
угри (Electrophorus electricus) и электрические скаты (Torpedo
spp.) производят импульсы, достаточно сильные, чтобы оглушить добычу (Sillar
et al. 2016) или нанести серьёзный вред людям или вызвать их смерть (Copenhaver
1991; Carlson 2015).
Де Янг (DeYoung 2000) и Гилмер (Gilmer 2011) высказали мысль о том, что динозавры
обладали электрическим органом, как у электрического угря, и использовали его
для образования искры, чтобы поджечь метан, произведённый в результате обмена
веществ. Однако, как мы уже увидели, воспламенение отрыгнутого метана окутало
бы голову животного огненным шаром. Вдобавок к этому, электрические органы не
образуют искры.
Искра – это электрический разряд в воздух. Диэлектрическая проницаемость (мера
лёгкости прохождения электрического тока сквозь материал) как у воздуха, так
и у метана низкая, около 1.0 (Wohlfarth 2013), тогда как у воды она – более
60 (Harvey 2013). По этой причине электрический ток, генерируемый электрической
рыбой, легко распространяется по воде или биологическим тканям (которые состоят
главным образом из воды), но не по воздуху или метану. Поскольку электрический
ток течёт по пути наименьшего сопротивления, ток, распространяющийся из одной
части тела животного в другую, будет протекать сквозь ткани животного, и не
будет проскакивать в виде искры через заполненный воздухом или метаном промежуток
между частями тела. Например, если у динозавра возникнет достаточное напряжение
между верхней и нижней челюстями, чтобы электрический ток протекал от одной
челюсти к другой, и динозавр откроет рот и отрыгнёт немного метана, сквозь облако
метана не проскочит ни одной искры; вместо этого ток пошёл бы из одной челюсти
в другую сквозь челюстные мускулы динозавра. Не было бы ни искры, ни пламени,
ни любого другого видимого свидетельства того, что имело место событие электрической
природы (рисунок 1). Поэтому данная гипотеза образования огня фальсифицирована.
|
Воспламенение горючего вещества искрой, образуемой путём трения
Гилмер (Gilmer 2011) выдвинул гипотезу, согласно которой динозавр мог использовать трение, чтобы высечь искру для поджигания горючего газа вроде метана «во рту, в горле, [или] во внутренних органах». Однако никто из животных не выделяет вещество, которое образует искры в ответ на трение. Твёрдые материалы, которые производятся телами животных – фосфат кальция, карбонат кальция, хитин и кератинизированные производные покровов тела – не дают искр, если тереть их друг об друга. Кремень, дающий искры, если тереть друг об друга его кусочки, является формой кварца (SiO2), химического соединения, которое могут осаждать некоторые микроорганизмы (Erlich and Newman 2009). Однако даже если бы осаждающие кремень микроорганизмы населяли рот или горло динозавров, любой метан, подожжённый там, взорвался бы там же, вызывая серьёзные внутренние повреждения.
Внутренние органы – бескислородная среда, поэтому они не позволят пламени появиться. Единственное исключение из этого правила – дыхательные пути, но пламя, образующееся здесь, вызывает серьёзные повреждения или смерть (Wöllmer et al. 2010). Даже в пищеварительном тракте находится слишком мало газообразного O2 для поддержания пламени (Levy 1954; Cunha et al. 2011) (рисунок 1). Поэтому данная гипотеза оказывается фальсифицированной.
Испускание самовоспламеняющейся пары химических соединений
Айзекс (Isaacs 2010), Бэтдорф и Порш (Batdorf and Porch 2013),
и Лейси (Lacy 2013) выдвинули гипотезу, согласно которой динозавры могли образовывать
огонь путём выделения пары химических соединений, которые загорелись бы в воздухе
после контакта друг с другом, будучи распылёнными изо рта или носа. Пара химических
соединений, спонтанно возгорающихся при смешивании, без стороннего источника
воспламенения, называется гипергольной. В гипергольную пару химических
соединений входят соединение-горючее и соединение-окислитель. Известно множество
гипергольных пар химических веществ, но в большинстве своём такие соединения
созданы человеком, и либо не встречаются в природе, либо, как в случае жидкого
кислорода, должны быть охлаждены до температур, которые не могут выдержать тела
животных. Два исключения – пероксид водорода (окислитель) и аммиак (горючее):
они оба производятся живыми организмами. Однако те виды горючего, с которыми
пероксид водорода образует гипергольную смесь – керосин, пентаборан, или смеси,
включающие гидразин и метанол (Sutton 2006) – чрезвычайно ядовиты для живых
организмов. Аналогичным образом окислители, с которыми образует гипергольную
смесь аммиак – жидкий кислород и жидкий фтор (Sutton 2006) – обладают температурой
кипения, слишком низкой (-183 ºC и -188 ºC соответственно [Compressed Gas Association
1990; Hammond 2013]) для того, чтобы живые организмы могли выдержать их присутствие
в жидком виде. Поэтому не существует ни одной известной гипергольной пары химических
соединений, выделение которых способны выдержать живые организмы.
Кроме того, если бы одно или оба вещества были газообразными, выброс гипергольной
пары химических соединений нанёс бы вред животному. Поскольку сразу после выпуска
из ёмкости газ рассеивается во всех направлениях, пара газообразных гипергольных
химикатов окружила бы голову динозавра, которая оказалась бы внутри огненного
шара, когда они прореагируют друг с другом. Если бы один из компонентов смеси
был газообразным, а другой – жидким, то после выброса газообразный компонент
сразу же распространится во всех направлениях, и достигнет места, из которого
жидкий компонент выходит из тела, нанося животному ожог в этом месте. Поэтому
данные варианты гипотезы самовоспламеняющейся смеси фальсифицированы.
Если бы оба химических соединения были жидкостями, их следовало бы распылять под такими углами, чтобы эти две струи соединялись достаточно далеко от животного, чтобы не обжечь его. Однако данная версия гипотезы будет, тем не менее, фальсифицирована отсутствием какой-либо пары гипергольных химических веществ, присутствие обоих компонентов которой способны выдержать тела животных.
Гребни динозавров-ламбеозаврин
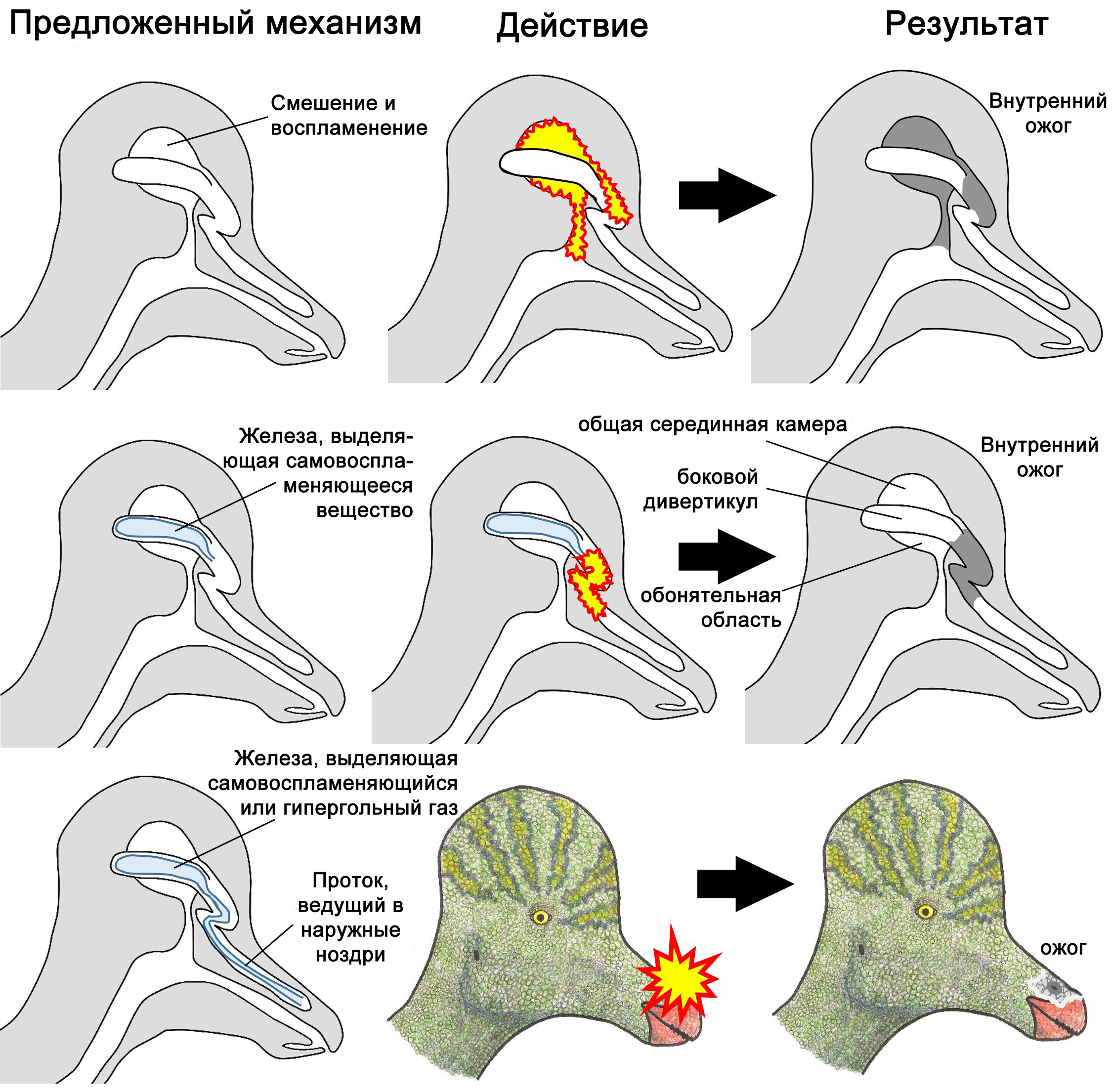
Утконосые динозавры подсемейства Lambeosaurinae обладали полыми гребнями, и в нескольких публикациях МЗК, в том числе в учебниках биологии для седьмого класса, выпущенных издательством BJU Press (Batdorf and Porch 2013; Lacy 2013), эти гребни интерпретируются как камеры для хранения или смешивания горючих газов (Gish 1977; Petersen 1986; Morris 1988; Niermann 1994; DeYoung 2000; Petersen 2002). Однако полые ходы в гребнях – часть дыхательных путей (Evans и др. 2009). Нечто запасаемое или смешиваемое там затруднило бы прохождение воздуха, вызывая удушье. Кроме того, левые и правые ходы разделены, что исключает возможность смешивания, кроме как в самом заднем отделе, где располагается обонятельный эпителий (Evans и др. 2009). Нечто, смешиваемое и воспламеняющееся там, вдобавок к причинению внутренних ожогов уничтожит обоняние у животного (рисунок 2).
|
В черепе утконосых динозавров подсемейства ламбеозаврин имеются боковые дивертикулы (ответвления) – слепо замкнутые мешки, присоединённые к левому и правому носовым ходам и тянущиеся вверх от носовой полости (Evans и др. 2009) (рисунок 2). Дивертикул лежит латеральнее по отношению к воздушному ходу, поэтому железа или приспособление для запасания вещества, расположенное там, не затруднили бы прохождения воздуха. Однако у архозавров наших дней (крокодилов и птиц) дивертикулы носовых проходов содержат лишь воздух, но не железы или приспособления для запасания веществ (Witmer 1997; Witmer и Ridgely 2008). Кроме того, методы зажигания огня, задействующие боковые дивертикулы ламбеозаврин, нанесли бы животному вред. Если в боковых дивертикулах размещались железы, которые выпускали пирофорные химические соединения в воздух внутри дыхательных путей, это вызвало бы возгорание внутри гребня (рисунок 2), вызывая внутренние ожоги. Если проток выводил пирофорные соединения к ноздре, гарантируя, что возгорание произойдёт вне гребня, ткань вокруг ноздри пострадает от ожогов (рисунок 2). Если в каждом из боковых дивертикулов находилась железа, которая выделяла один из газов гипергольной пары, то после выхода из ноздрей газы рассеются во всех направлениях и воспламенятся, охватывая морду животного (или, возможно, всю голову) огненным шаром.
Если в каждом из боковых дивертикулов находилась железа, которая выделяла один компонент из гипергольной пары жидкостей, а не газов, и если протоки выводили жидкости в ноздри, а жидкости выбрызгивались сжатием мускулатуры, и две струи жидкости сливались вместе достаточно далеко от морды, чтобы избежать нанесения ожогов, когда эти две струи контактируют друг с другом и загораются, то животное могло бы избежать повреждений. Однако в черепе у ламбеозаврин на костях полностью отсутствуют признаки присутствия необходимых желёз, протоков или мускулов (Evans и др. 2009). Кроме того, не существует такой гипергольной пары химических соединений, синтез обоих компонентов которой способны выдержать тела животных. Поэтому данная гипотеза фальсифицирована.
Хрящевые подрывные капсюли?
Айзекс (Isaacs 2010) выдвинул гипотезу, согласно которой у динозавров был «кусок хряща», который продолжался за пределы образованной костями части рыла, как у млекопитающих, и в нём, «возможно, находилось место для смешивания химических соединений и кислорода, используемых для горения». Однако у известных нам животных носовые хрящи не вмещают в себя приспособления для зажигания огня. И что гораздо важнее, если бы внутри хрящевого «куска» происходило горение, эта структура пострадала бы от ожогов, что фальсифицирует эту гипотезу.
Прочие мезозойские рептилии
|
Букер (Booker 2005) выдвинул гипотезу о том, что увеличенная полость на конце морды у мелового крокодиломорфа Sarcosuchus вмещала источник огня. Виланд (Wieland 2005) и Пол (Paul 2010) предположили, что полость использовалась для смешивания горючих газов. В учебниках биологии для седьмого класса Бэтдорф и Порш (Batdorf and Porch 2013) и Лейси (Lacy 2013) также предполагали, что полость использовалась для зажигания огня.
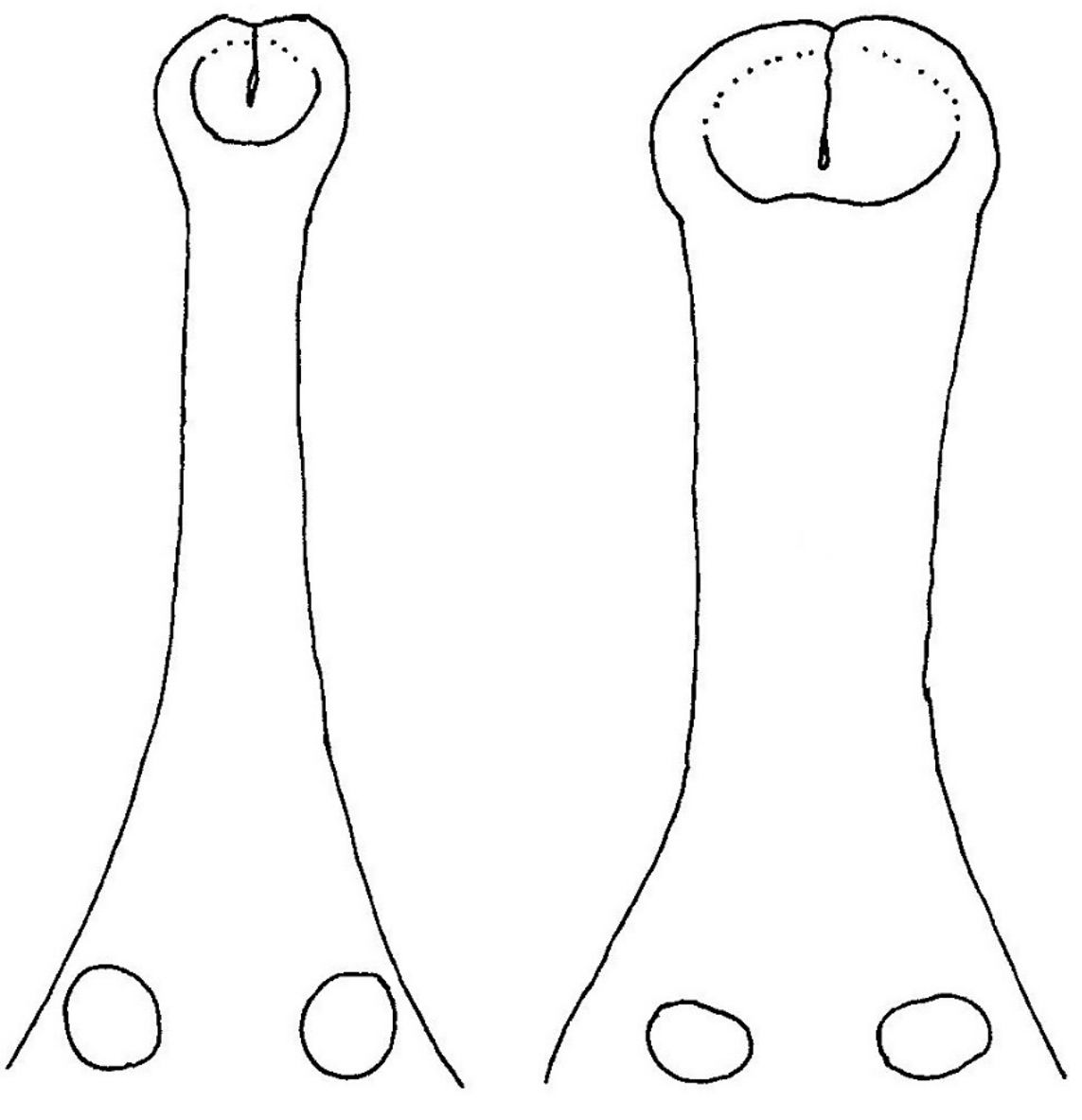
Появление увеличенной полости на конце морды у взрослого Sarcosuchus происходит в результате расширения морды в процессе индивидуального развития. У молодых особей Sarcosuchus узкие морды, и в процессе индивидуального развития морда расширяется по всей длине, потому кончик морды расширяется не отдельно, а вместе с остальной частью морды (Sereno и др. 2001) (рисунок 3). Поэтому широкий кончик морды взрослой особи связан с развитием широкой морды в целом, а не с неким особым назначением кончика морды. Узкомордые крокодилы питаются почти исключительно рыбой, тогда как виды со средней и большой шириной морды – неспециализированные хищники с рационом, который включает крупную добычу (McHenry и др. 2006). Поэтому изменение в размерах морды у Sarcosuchus функционально связано с изменением рациона по мере роста животного.
Расширение морды в сочетании с отсутствием окостеневшей интернариальной перегородки между ноздрями создаёт иллюзию увеличенной полости на кончике морды Sarcosuchus. У многих ныне живущих видов крокодилов также отсутствует окостеневшая интернариальная перегородка, а вместо неё имеется перегородка из мягких тканей, которая разделяет ноздри (Iordansky 1973). Нет никаких причин подозревать, что у Sarcosuchus всё было иначе, поэтому наличие у животного во плоти единственной увеличенной полости очень маловероятно. Даже если бы она существовала, разжигание огня внутри неё вызвало бы появление ожога у животного, что фальсифицирует эту гипотезу.
Гилмер (Gilmer 2011) выдвинул гипотезу, согласно которой гребни «некоторых птерозавров вроде Pteranodon» «могли содержать воспламеняющийся газ… для использования при необходимости». Эта гипотеза анатомически нереалистична; у некоторых птерозавров, обладающих гребнями, возможно, что внутри гребня находились наполненные воздухом ответвления носовой полости или полости среднего уха (Bennett 2001), но гребни чрезвычайно тонкие на поперечном срезе, и в них недостаточно места для запасания чего-либо, и у ныне живущих архозавров носовые ответвления содержат только воздух (Witmer 1997; Witmer and Ridgely 2008). Кроме того, у самого рода Pteranodon отсутствуют признаки того, что лобная кость, которая образует весь гребень, пронизана выростами носовой полости, что самое главное (Bennett 2001). Поэтому данная гипотеза фальсифицирована.
Истинное происхождение легенд о драконах
Биологическая действительность, стоящая за происхождением легенд
о драконах, известна уже очень давно. Слово «дракон» происходит от древнегреческого
δράκων (drakōn) и латинского draco – они оба означают «змей». Многочисленные
древние греческие и римские артефакты иллюстрируют мифы о drakōn/draco,
и на них на всех изображённое животное – змея (Ogden 2013; Senter 2013). Греческие
и римские работы по естествознанию описывали drakōn/draco как змею.
Некоторые из таких работ ограничивались применением этого термина к крупным
неядовитым змеям-душителям, в частности, к эскулапову полозу (Zamenis longissima),
или к африканским и индийским питонам (Senter 2013; Senter и др. 2016).
Европейские знания о драконах эволюционировали в Средние Века. Слухи о том,
что драконы могли летать и метать огонь, начались с пятого века (Senter и др.
2016). К десятому веку драконы обычно изображались с оперёнными крыльями и парой
конечностей (Temple 1976; Mittman 2006). Описание драконов как четвероногих
животных с крыльями как у летучей мыши началось с тринадцатого века и стало
обычным в эпоху Возрождения (Allen и Griffiths 1979; Benton 1992; Absalon и
Canard 2006; Morrison 2007). К девятнадцатому веку, когда натуралисты дали динозаврам
их первые научные описания, европейские художники на протяжении уже примерно
четырёх веков регулярно изображали драконов, имеющих до странности динозавровую
или птерозавровую внешность. Естественно, за этим последовали предположения
о том, что встречи людей с этими животными стали источником вдохновения для
легенд о драконах, и литература МЗК в наше время обычно изображает динозавров
как огнедышащих существ. Но, как было показано здесь, эта точка зрения нереалистична,
и продолжать высказывать её в научных учебниках – это чистейшая безответственность.
• Absalon, Patrick, and Frédérik Canard. 2006. Les Dragons. Des Monstres
au Pays des Hommes. Paris: Gallimard.
• Allen, Judy, and Jeanne Griffiths. 1979. The Book of the Dragon.
London: Orbis.
• Aneshansley, Daniel J., Thomas Eisner, Joanne M. Widom, et al. 1969. Biochemistry
at 100° C: Explosive secretory discharge of bombardier beetles (Brachinus).
Science 165: 61–63.
• Batdorf, Brad R., and Thomas Porch. 2013. Life Science, 3rd edition.
Greenville, SC: BJU Press.
• Bennett, S. Christopher. 2001. The osteology and functional morphology
of the Late Cretaceous pterosaur Pteranodon. Palaeontographica Abteilung
A 260: 1–112.
• Benton, Janetta R. 1992. The Medieval Menagerie. Animals in the Art of
the Middle Ages. New York: Abbeville Press.
• Booker, Peter. 2005. A new candidate for Leviathan? Technical Journal
19: 14–16.
• Carlson, Bruce A. 2015. Animal behavior: Electric eels amp up for an easy
meal. Current Biology 25: R1070–R1072.
• Colvin, H.W., P.T. Cupps, and H.H. Cole. 1958. Dietary influences on eructation
and related ruminal phenomena in cattle. Journal of Dairy Science 41: 1565–1579.
• Compressed Gas Association. 1990. Handbook of Compressed Gases. New
York: Chapman and Hall.
• Copenhaver, Brian P. 1991. A tale of two fishes: Magical objects in natural
history from antiquity through the scientific revolution. Journal of the
History of Ideas 52: 373–398.
• Cunha, Isabel S., Cristine C. Barreto, Ohana Y.A. Costa, et al. 2011. Bacteria
and Archaea community structure in the rumen microbiome of goats (Caphampra
hircus) from the semiarid region of Brazil. Anaerobe 17: 118–124.
• DeYoung, Donald B. 2000. Dinosaurs and Creation. Grand Rapids: Baker.
• Erlich, Henry L., and Dianne K. Newman. 2009. Geomicrobiology, 5th
Edition. Boca Raton: CRC.
• Evans, David C., Ryan Ridgely, and Lawrence M. Witmer. 2009. Endocranial
anatomy of lambeosaurine hadrosaurids (Dinosauria: Ornisthichia): A sensorineural
perspective on cranial crest function. Anatomical Record 292: 1315–1337.
• Gallant, Jason R., Lindsay L. Traeger, Jeremy D. Volkening, et al. 2014. Genomic
basis for the convergent evolution of electric organs. Science 344: 1522–1525.
• Galley, Archibald H. 1954. Combustible gases generated in the alimentary
tract and other hollow viscera and their relationship to explosions occurring
during anaesthesia. British Journal of Anaesthesia 26: 189–193.
• Gilmer, James E. 2011. 100 Year Cover-up Revealed. We Lived with Dinosaurs.
Bloomington: Authorhouse.
• Gish, Duane T. 1977. Dinosaurs, Those Terrible Lizards. San Diego:
Creation-Life Publishers.
• Gradstein, Felix, James Ogg, and Alan Smith. 2004. A Geologic Time Scale
2004. Cambridge: Cambridge University Press.
• Haddock, Steven H.D., Mark A. Moline, and James F. Case. 2010. Bioluminescence
in the sea. Annual Review of Marine Science 2: 442–493.
• Hammond, C.R. 2013. The elements. In William M. Haynes, David R.
Lide, and Thomas J. Bruno. (eds.), CRC Handbook of Chemistry and Physics,
94th Edition. Boca Raton: CRC, 4.1–4.42.
• Hamp, Douglas. 2000. The First Six Days. Confronting the God-plus-evolution
Myth. Santa Ana: Yoel.
• Harvey, Allan H. 2013. Permittivity (dielectric constant) of water at
various frequencies. In William M. Haynes, David R. Lide, and Thomas J.
Bruno. (eds.), CRC Handbook of Chemistry and Physics, 94th Edition.
Boca Raton: CRC, 6.14.
• Iordansky, Nikolai N. 1973. The skull of the Crocodilia. In Carl
Gans and Thomas S. Parsons (eds.), Biology of the Reptilia, Volume
4: Morphology D. London: Academic, 201–262.
• Isaacs, Darek. 2010. Dragons or Dinosaurs? Creation or Evolution?
Alachua, FL: Bridge-Logos.
• Koch, A.K.S., P. Nørgaard, and K. Hilden. 2009. A new method for simultaneous
recording of methane eructation, reticulo-rumen motility and jaw movements in
rumen fistulated cattle. In Y. Chilliard, F. Glasser, Y. Faulconnier, et
al. (eds.), Ruminant Physiology. Digestion, Metabolism, and Effects of Nutrition
on Reproduction and Welfare. Wageningen, Netherlands: Wageningen Academic,
360–361.
• Lacy, Elizabeth A. 2013. Life Science, 4th Edition. Greenville, SC:
BJU Press.
• Levy, Edward I. 1954. Explosions during lower bowel electrosurgery. A
method of prevention. American Journal of Surgery 88: 754–758.
• Lissmann, H.W. 1958. On the function and evolution of electric organs
in fish. Journal of Experimental Biology 35: 156–191.
• MacDonald, A.G. 1994. A brief historical review of non-anaesthetic causes
of fires and explosions in the operating room. British Journal of Anaesthesia
73: 847–856.
• McHenry, Colin R., Philip D. Clausen, William J.T. Daniel, et al. 2006. Biomechanics
of the rostrum in crocodilians: A comparative analysis using finite-element
modeling. Anatomical Record Part A 288A: 827–849.
• Mittman, Asa. 2006. Maps and Monsters in Medieval England. New York:
Routledge.
• Morris, Henry M. 1984. The Biblical Basis for Modern Science. Grand
Rapids: Baker.
• ———. 1988. The Remarkable Record of Job. Grand Rapids: Baker.
• Morris, John D. 1999. Dinosaurs, the Lost World, and You. Green Forest,
AR: Master Books.
• Morrison, Elizabeth. 2007. Beasts Factual and Fantastic. The Medieval
Imagination. Los Angeles: J. Paul Getty Museum.
• Niermann, D. Lee. 1994. Dinosaurs and dragons. Creation Ex Nihilo
Technical Journal 8(1): 85–104.
• Ogden Daniel. 2013. Drakōn: Dragon Myth and Serpent Cult in the Greek
and Roman Worlds. Oxford: Oxford University Press.
• Paul, Mart-Jan. 2010. Behemoth and Leviathan in the book of Job. Journal
of Creation 24(3): 94–100.
• Petersen, Dennis R. 1986. Unlocking the Mysteries of Creation. El
Dorado, CA: Creation Resource Foundation.
• ———. 2002. Unlocking the Mysteries of Creation, Premier Edition.
Alachua, FL: Bridge-Logos.
• Phillips, Phil. 1994. Dinosaurs, the Bible, Barney and Beyond. Lancaster,
PA: Starburst.
• Pough, F. Harvey, Christine M. Janis, and John B. Heiser. 2013. Vertebrate
Life. 9th Edition. Boston: Pearson.
• Prothero, Donald. 2007. Evolution. What the Fossils Say and Why It Matters.
New York: Columbia University Press.
• Senter, Phil. 2013. Dinosaurs and pterosaurs in Greek and Roman art and
literature? An investigation of young-Earth creationist claims. Palaeontologia
Electronica 16(3.25A): 1–16.
• Senter, Phil, Uta Mattox, and Eid E. Haddad. 2016. Snake to monster: Conrad
Gessner’s Schlangenbuch and the evolution of the dragon in the literature of
natural history. Journal of Folklore Research 53: 67–124.
• Sereno, Paul C., Hans C.E. Larsson, Christian A. Sidor, et al. 2001. The
giant crocodyliform Sarcosuchus from the Cretaceous of Africa. Science
294: 1516–1519.
• Sillar, Keith, Laurence D. Picton, and William J. Heilter. 2016. The Neuroethology
of Predation and Escape. Oxford: John Wiley and Sons.
• Sutton, George P. 2006. History of Liquid Propellant Rocket Engines.
Reston, VA: American Institute of Aeronautics and Astronautics.
• Temple, Elżbieta. 1976. Anglo-Saxon Manuscripts 900–1066. London:
Harvey Miller.
• Tsai, Hsiao-Yun, Sheng-Wei Wang, Sin-Ying Wu, et al. 2010. Experimental
studies on the ignition behavior of pure silane released into air. Journal
of Loss Prevention in the Process Industries 23: 170–177.
• Ulyatt, M.J., S.K Baker, G.J. McCrabb, et al. 1999. Accuracy of SF6
tracer technology and alternatives for field measurements. Australian Journal
of Agricultural Research 50: 1329–1334.
• Wieland, Carl. 2005. Dragons of the Deep. Green Forest, AR: Master
Books.
• Witmer, Lawrence M. 1997. The evolution of the antorbital cavity of archosaurs:
A study in soft-tissue reconstruction in the fossil record with an analysis
of the function of pneumaticity. Society of Vertebrate Paleontology Memoir
3: 1–73.
• Witmer, Lawrence M., and Ryan C. Ridgely. 2008. The paranasal air sinuses
of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria)
and their contribution to cephalic structure. Anatomical Record 291: 1362–1388.
• Wohlfarth, Christian. 2013. Permittivity (dielectric constant) of gases.
In William M. Haynes, David R. Lide, and Thomas J. Bruno. (eds.), CRC Handbook
of Chemistry and Physics, 94th Editon. Boca Raton: CRC, 6.209.
• Wöllmer, Wolfgang, Götz Schade, and Gerhard Kessler. 2010. Endotracheal
tube fires still happen—a short overview. Medical Laser Application 25:
118–125.
Филип Дж. Сентер – специалист по палеонтологии позвоночных со специализацией в области палеобиологии динозавров. Он получил степень доктора философии в области биологических наук в Университете Северного Иллинойса в 2003 году, и ведёт курсы биологии в Государственном университете Фейетвилла в Северной Каролине. Он опубликовал более шестидесяти статей о палеонтологии динозавров, биологии рептилий и дебатах сторонников творения и эволюции. С ним можно переписываться по адресу psenter@uncfsu.edu
Заявление Филипа Сентера о науке и Церкви:
Я, Филип Сентер, не только учёный, но также греческий православный
христианин. В дополнение к степени доктора философии в биологических науках
у меня есть степень магистра богословия в православном христианстве. Как учёный,
я соглашаюсь с полученными в природе данными, которые показывают, что Земле
миллиарды лет, и что все организмы на Земле эволюционировали от общего предка.
Как христианин и библеист я согласен с многочисленными эпизодами в Библии, которые
учат нас, что Бытие и остальная часть Пятикнижия – это не исторические сведения,
и их не следует толковать буквально.
Как учит сама Библия, отдых Бога на седьмой день (Бытие 2:2) был не событием,
произошедшим в буквальный день в прошлом, а духовным состоянием покоя, которого
может достичь верующий (Евреи 3:7 – 4:11); двойственная природа Адама (частично
земля, частично дыхание жизни: Бытие 2:7) – это метафора двойственной природы
человека (частично материальная, частично духовная) (Первое послание к коринфянам
15:44-49), союз Адама и Евы олицетворяет отношения между Богом и людьми (К Эфесянам
5:31-33), Всемирный потоп – символ духовного очищения сердца (Первое послание
Петра 3:20-21), обещание Бога Аврааму о его семени относится к потомству в переносном,
а не в буквальном смысле (Послание к Римлянам 9:6-8; Послание к Галатам 3:7,
3:16, 3:29), история о рождении Исаака является аллегорией о потомстве в переносном
смысле (Послание к Римлянам 9:6-8; Послание к Галатам 4:22-31), предписание
о проведении обрезания относится к духовному, а не к физическому действию (Послание
к Римлянам 2:29), иные предписания о ритуалах из Пятикнижия – символические
отсылки к тайне Спасителя (Послание к Колоссянам 2:16-17; Послание к Евреям
9:7-9), различные события в истории пути Израильтян в Землю Обетованную – образные
отсылки к тайне спасения через Спасителя (Первое послание к Коринфянам 10:1-11),
история исхода и пути Израильтян к Земле Обетованной – это притча (Псалом 78:2),
а вход израильтян в Землю Обетованную покоя – метафорическая отсылка к обещанию
каждому верующему, что его ожидает состояние духовного покоя (Послание к Евреям
3:7 – 4:11). Через эти и другие отрывки сама Библия даёт христианам позволение
принять через природу божественное откровение о биологической эволюции и старой
Земле, поскольку эти отрывки показывают, что библейская книга, которая описывает
сотворение за шесть дней 6000 лет назад (Бытие), не предполагает её буквального
понимания.
Так что же представляют собой Книга Бытия и остальная часть Пятикнижия, если
это не собрание исторических записей? Согласно Отцам Церкви, эти книги – притчи:
теологические утверждения в форме историй. Они написаны символическим языком,
и должны пониматься в ином смысле, нежели буквально (примеры: св. Григорий Нисский,
«Большой Катехизис» 5 и 8; Климент Александрийский, «Строматы» 1.28; Тертуллиан
«Против Маркиона» 3.3; Аноним, «Послание Варнавы», 6.10). Несколько Отцов Церкви
даже написали трактаты о том, как интерпретировать символический язык в Книге
Бытия и остальном Пятикнижии (примеры: св. Григорий Нисский, «Жизнь Моисея»,
книга 2; св. Максим Исповедник, «Вопросоответы к Фалассию»; прп. Анастасий Синаит,
«Шестоднев»; Аноним, «Послание Варнавы»; св. Амвросий Mедиоланский, «Об Аврааме»;
Дидим Слепец, «На Бытие»; Тихоний Африканский, «Книга о семи правилах»; св.
Иустин Мученик, «Диалог с Трифоном иудеем»). Даже в рамках Библии св. Павел
характеризует священное писание Ветхого Завета как отправную точку для теологического
образования и исправления поведения (Второе послание к Тимофею 3:16), но не
называет его историческими записями.
Библия и Отцы Церкви, указанные в вышеприведённом списке, согласны с тем, что
Книга Бытия и остальное Пятикнижие не должны пониматься буквально. Поэтому верующий
может свободно принять выводы науки относительно биологической эволюции и старой
Земли. Важно знать, что человек, который принимает Церковь, может также принять
и науку.
| Главная |