|
| Главная | Неоцен |
|
SYLLOGEUS - издание Национального Музея Естествознания, Национальные Музеи Канады, предназначенное для облегчения быстрого распространения информации, относящейся к научным дисциплинам и просветительским функциям, которые находятся в ведении Национального музея естествознания. Ради того, чтобы быстро донести информацию, были ускорены обычные процедуры, связанные с публикацией.
Статьи публикуются на английском, французском, или на обоих языках, а выпуски издаются нерегулярно. Полный список названий изданий, выпущенных с момента основания серии (1972 г.), и отдельные копии этого выпуска доступны по почте от Национального Музея Естествознания, Оттава, Канада. KlA 0M8
Серия «Syllogeus», выпуск 37
(c) Национальные Музеи Канады, 1982 г.
Напечатано в КанадеISSN 0704-576X
РЕКОНСТРУКЦИИ МЕЛКОГО МЕЛОВОГО ТЕРОПОДА
Stenonychosaurus inequalis И ГИПОТЕТИЧЕСКОГО ДИНОЗАВРОИДА
Д. Расселл и Р. Сегуин
Национальный Музей Естествознания
Оттава K1A OM8
«Syllogeus» выпуск 37
Национальные Музеи Канады
Национальный Музей Естествознания
Дейнонихозавры – это мелкие плотоядные динозавры, которые в настоящее время разделены на две родословных линии: дромеозавры и более грацильные заврорнитоиды. Самый полный образец заврорнитоида, известный из Северной Америки - Stenonychosaurus inequalis из позднемеловых отложений близ Провинциального парка Дайносор в Альберте. Он был использован в качестве основы для создания раскрашенной модели этого вида в натуральную величину. Были высказаны предположения о том, как могли бы измениться потомки S. inequalis с его крупным мозгом, стереоскопическим зрением, противопоставляющимися пальцами и двуногим положением тела, если бы они пережили вымирание в конце мезозоя и достигли значения коэффициента энцефализации, подобного таковому у Homo sapiens. Модель этого гипотетического существа, динозавроида, была «реконструирована» и отлита в таком же стиле, как Stenonychosaurus, и было дано объяснение физических особенностей модели динозавроида.
В прошлом десятилетии группа мелких хищных динозавров, которые жили почти в самом конце эры рептилий, стала объектом особого внимания со стороны палеонтологов. Реконструкции одного из видов (Deinonychus) распространились повсеместно в специальных и популярных изданиях (например, Ostrom 1969B, фронтиспис; 1978A, стр. 160-161), а наличие у другого вида (Stenonychosaurus) увеличенного мозга дало толчок спорам об эволюции разума у рептилий (например, Hopson 1980, Расселл 1981). Для последнего вида выполнено очень мало реконструкций, и здесь представлена новая, призванная частично удовлетворить необходимость в них. В дополнение к своим типично динозавровым признакам Stenonychosaurus, возможно, обладал также перекрывающимися полями зрения и противопоставляющимся пальцем в кисти. Это сочетание особенностей вызвало множество неофициальных вопросов о том, какой могла бы быть последующая история этих животных, если бы динозавры не вымерли. В качестве ответа на данные вопросы здесь представлена предположительная объёмная модель. Можно надеяться, что эта модель будет способствовать обсуждению влияния энцефализации на морфологию позвоночных.
Хотя наше профессиональное обучение проходило в различных областях (Д. Р. – палеонтология, Р. С. - таксидермия), две модели, обсуждаемые ниже – это результат совместной работы, в которой вклад каждого из нас сложно представить по отдельности.
Мы выражаем благодарность докторам Фрэнсису Куку и Хенри Уэллету из отдела зоологии позвоночных Национального музея естествознания за информацию о живорождении у рептилий и морфологии когтей у хищных птиц; доктору Филипу Курри из Провинциального Музея Альберты за слепок образца Stenonychosaurus из коллекции его учреждения; доктору Арнольду Вигерту из лаборатории гидравлики Национального научно-исследовательского совета Канады за измерение объёма воды, вытесненной готовой моделью Stenonychosaurus, и ещё нашим коллегам из отдела палеобиологии за многие часы продуктивных дискуссий в течение всего времени работы над моделями.
Мы хотим поблагодарить миссис Гейл Райс и доктора К. Р. Харингтона за помощь в подготовке окончательного черновика этой публикации. Мисс Нэнси Аршамбо (Systemhouse Ltd.) перепечатала рукопись.
Сокращения названий учреждений, которые указаны перед номерами упомянутых в тексте образцов, следующие:
AMNH - Американский музей естественной истории
NMC - Национальные музеи Канады
PMA - Провинциальный Музей Альберты
YPM – Музей Пибоди в Йельском университете
Примерно пятьдесят лет назад в позднемеловых отложениях в Северной Америке и Центральной Азии были идентифицированы останки нескольких мелких плотоядных динозавров, родственных друг другу (Таблица 1). Сообщество палеонтологов не оценило анатомическое своеобразие этих существ должным образом до тех пор, пока морфология Deinonychus – раннемелового представителя этой группы – не была подробно описана Остромом (Ostrom 1969A, 1969B, 1976). После этого они были признаны особым инфраотрядом динозавров-теропод, которые обладали хищническими когтями на внутренней стороне в остальном двупалой ступни, приспособленной для бега, и получили название дейнонихозавры (Colbert and Russell 1969, Ostrom 1969B). Назначение этих когтей как приспособления для вырывания внутренностей наглядно продемонстрировало открытие сохранившегося в ископаемом виде скелета Velociraptor, вцепившегося в тело мелкого растительноядного динозавра (Protoceratops, см. Barsbold 1974A; Ostrom 1978A, фотография на стр. II). О наличии хищнических когтей на внутренней стороне ступни также сообщалось в отношении Naosaurus, мелкого теропода из самого конца мела Аргентины (Bonaparte и Powell 1980)*. Если эти структуры не появились в процессе эволюции независимо, их можно приводить в качестве свидетельства того, что дейнонихозавры возникли и распространились по всему миру до разделения континентов в раннем мелу (см. также Galton 1980). Также были отмечены признаки сходства между скелетами Archaeopteryx, которого большинство учёных рассматривает как предковую позднеюрскую птицу, и дейнонихозавров (Ostrom 1974, Barsbold and Perle 1979). Однако ступня Archaeopteryx лишена признаков специализации, наблюдаемых у дейнонихозавров (см. также Wellnhofer 1974), и их родство не может быть близким.
|
В настоящее время среди Deinonychosauria выделены две группы: Dromaeosauridae, включающая Deinonychus, Velociraptor и Dromaeosaurus, и Saurornithoididae, включающая Saurornithoides и Stenonychosaurus (Ostrom 1969B, рис. 83; Colbert and Russell 1969, стр. 39; Russell 1969; Barsbold 1974B, 1976; Sues 1978). Общая морфология дромеозавров в настоящее время понята сравнительно хорошо и считается типичной для инфраотряда. Однако заврорнитоиды отличаются от дромеозавров по многим признакам скелета. Эти животные были более изящными, чем дромеозавры. Череп и зубы были относительно маленькими. Глаза и мозг были крупнее, чем у дромеозавров, а глаза были сдвинуты в антеромедиальное положение, что позволяет предположить наличие значительно перекрывающихся полей зрения. Тела предкрестцовых позвонков лишены полостей (плевроцелей), наблюдаемых у дромеозавров, а зигапофизы дистальных хвостовых позвонков короче. Таким образом, заврорнитоиды представляли собой самостоятельную группу позднемеловых теропод.
Самый полный образец из числа заврорнитоидов среди известных на настоящий момент из Северной Америки – NMC 12340, принадлежащий Stenonychosaurus inequalis (Russell 1969), который примерно так же полон, как известный скелет австралопитецина «Люси» из области Афар в Эфиопии (Johanson and Edey 1981). Он был собран в отложениях речной дельты (Джудит Ривер = формация Олдман) возрастом 76 миллионов лет близ Провинциального парка Дайносор в Альберте; обзор её палеоэкологии был сделан Беландом и Расселлом (Béland and Russell 1978). Размеры скелетных элементов, сохранившихся у NMC 12340, позволили сделать привязку по масштабу для трёхмерной пластиковой реконструкции скелета (Рис. 1-5), в которой была сведена воедино морфология известных скелетных фрагментов S. inequalis в общем соответствии с более полным материалом близкородственных форм.
Череп (рис. 6) смоделирован на основе фрагментов черепа, сохранившихся у NMC 12340, отдельной верхней челюсти NMC 12392, зубной кости NMC 8540 (см. также Russell 1969) и лобно-теменной кости, на которой исключительно хорошо сохранились гребни вдоль постериорного края височных ямок (PMA P79.8.1), с добавлением особенностей морфологии, сохранившихся на черепах Saurornithoides mongoliensis и S. junior (Barsbold 1974B). Если череп был реконструирован точно, он шире и короче, чем у азиатских видов (cf. Russell 1969, стр. 600).

РИСУНОК 1. Реконструированный скелет STENONYCHOSAURUS INEQUALIS,
вид сбоку (NMC neg. 81-1668).

РИСУНОК 2. Реконструированные череп и передняя часть осевого
скелета STENONYCHOSAURUS
INEQUALIS (NMC neg. 81-1671).
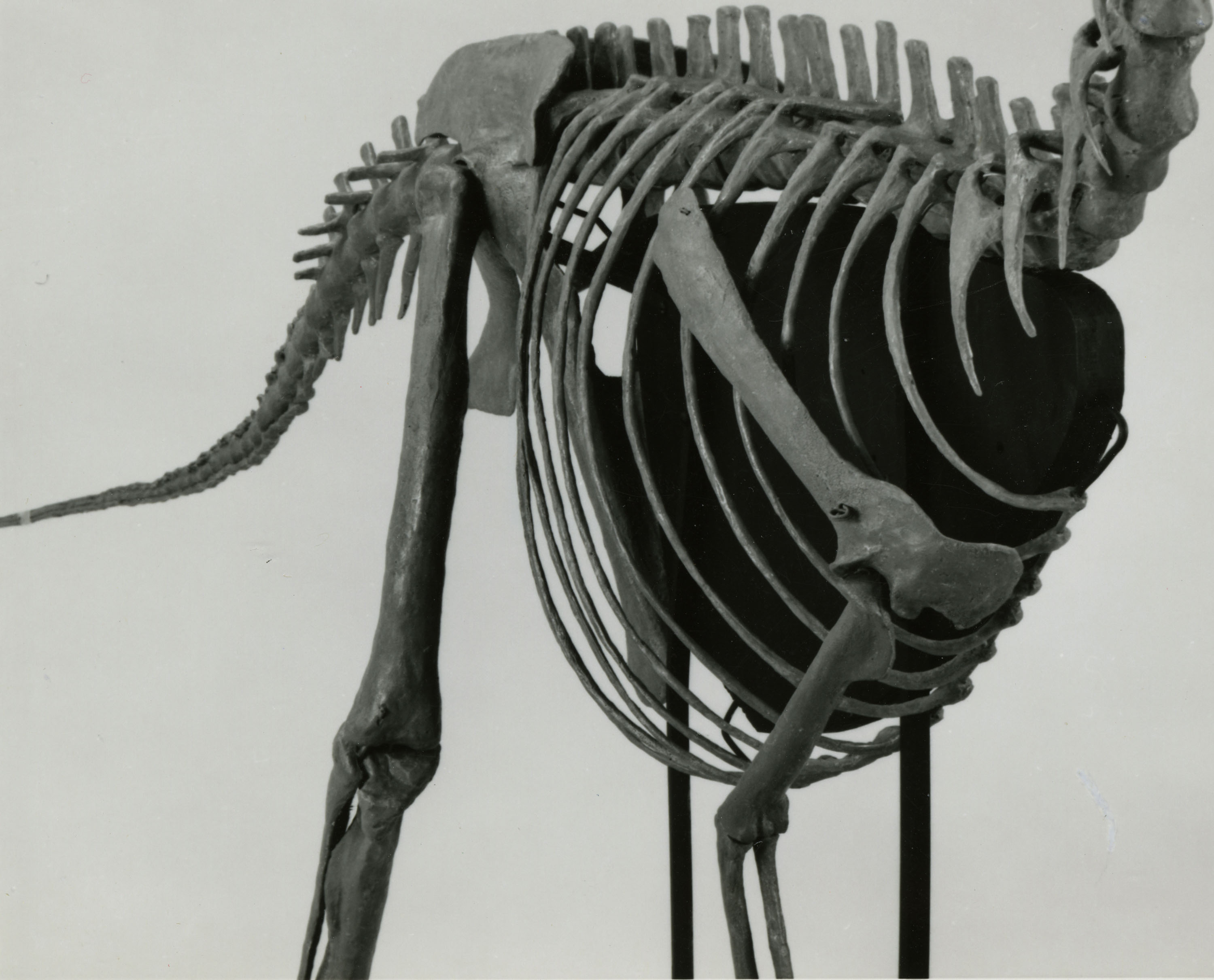
РИСУНОК 3. Реконструированные пояса конечностей и осевой скелет
STENONYCHOSAURUS INEQUALIS, вид спереди и сбоку (NMC neg. 81-1667).
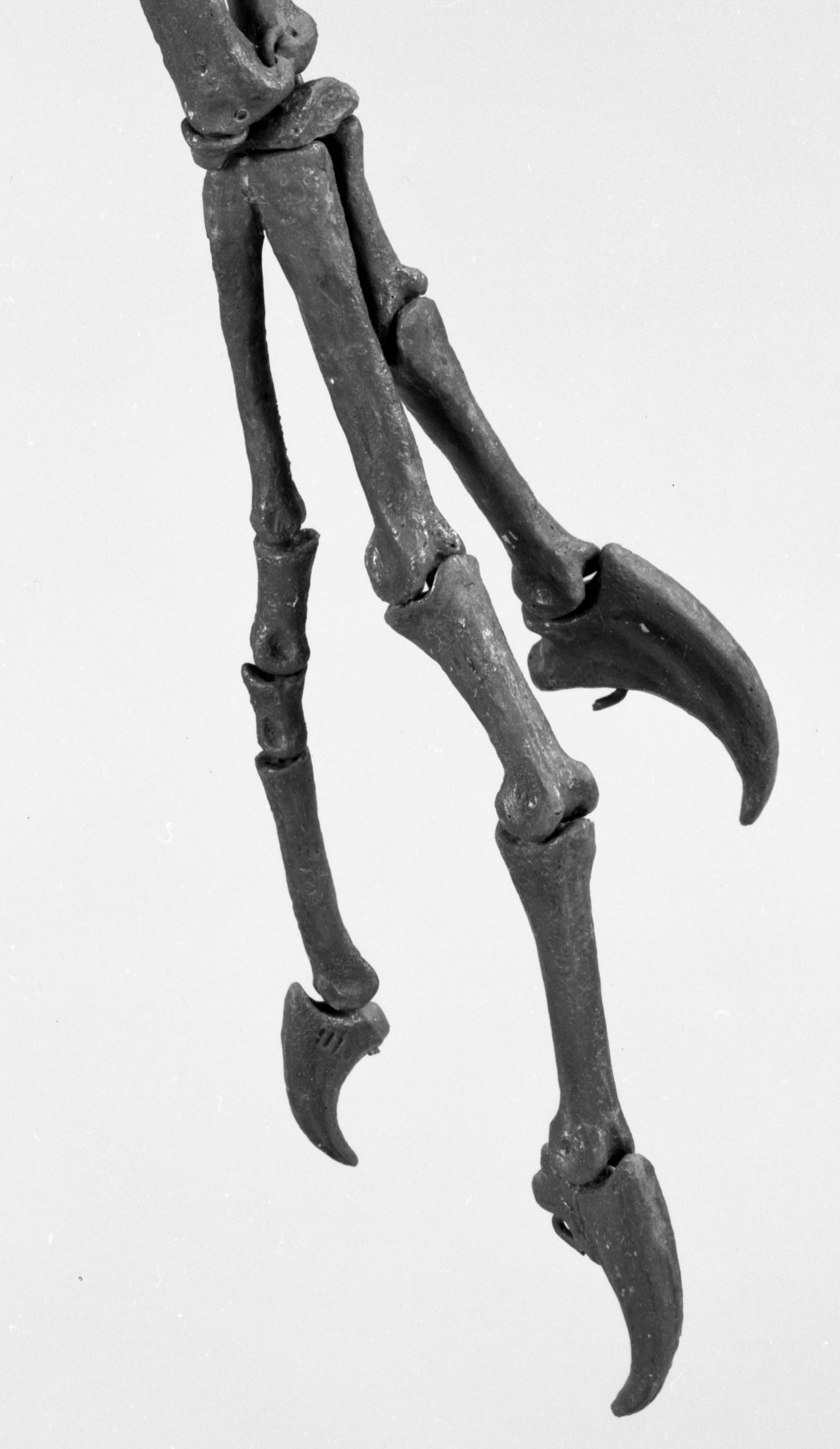
РИСУНОК 4. Реконструированная кисть STENONYCHOSAURUS INEQUALIS,
вид сверху
(NMC neg. 81-1670).
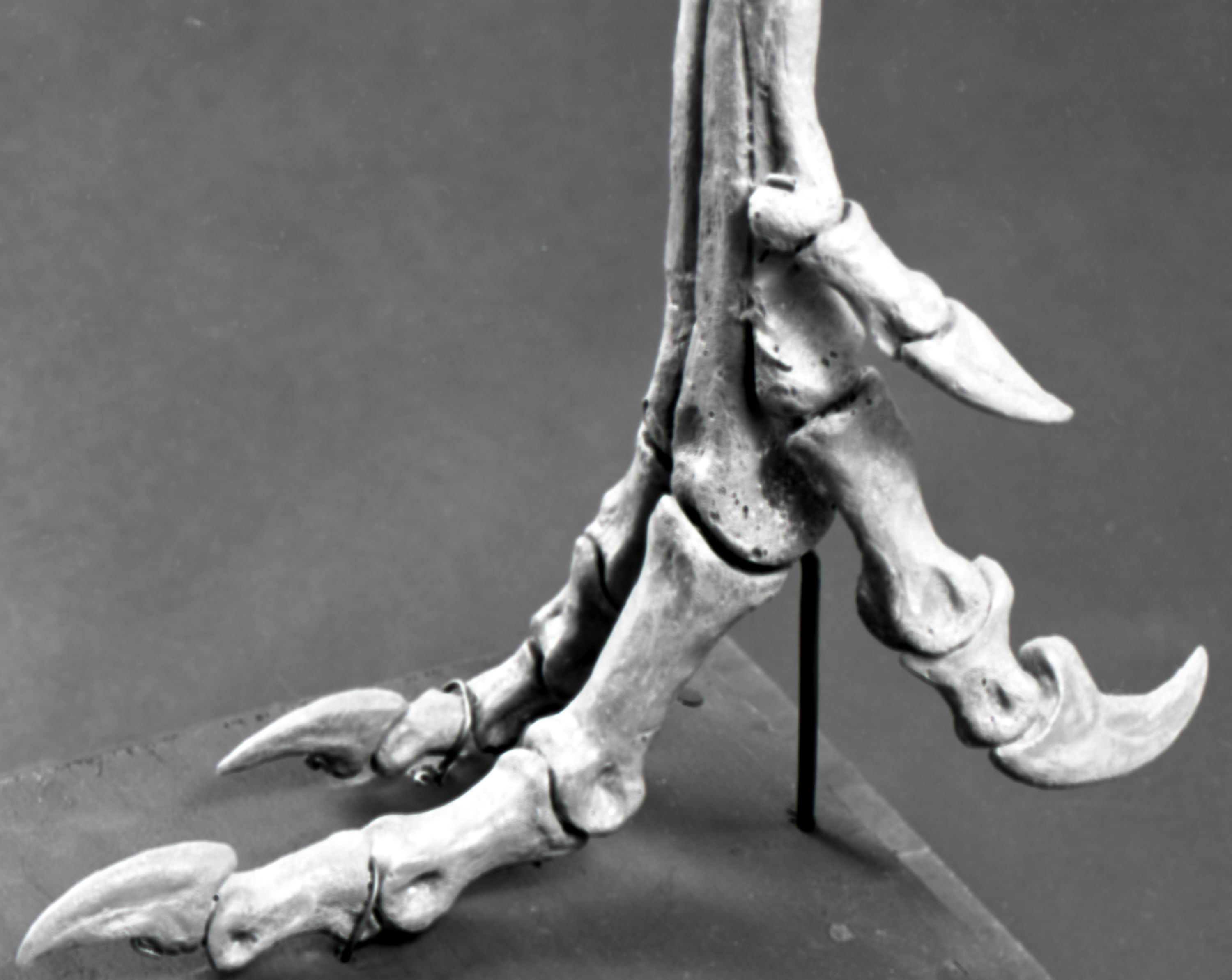
РИСУНОК 5. Реконструированная ступня STENONYCHOSAURUS INEQUALIS,
вид с медиальной стороны
(NMC neg. 81-1674).
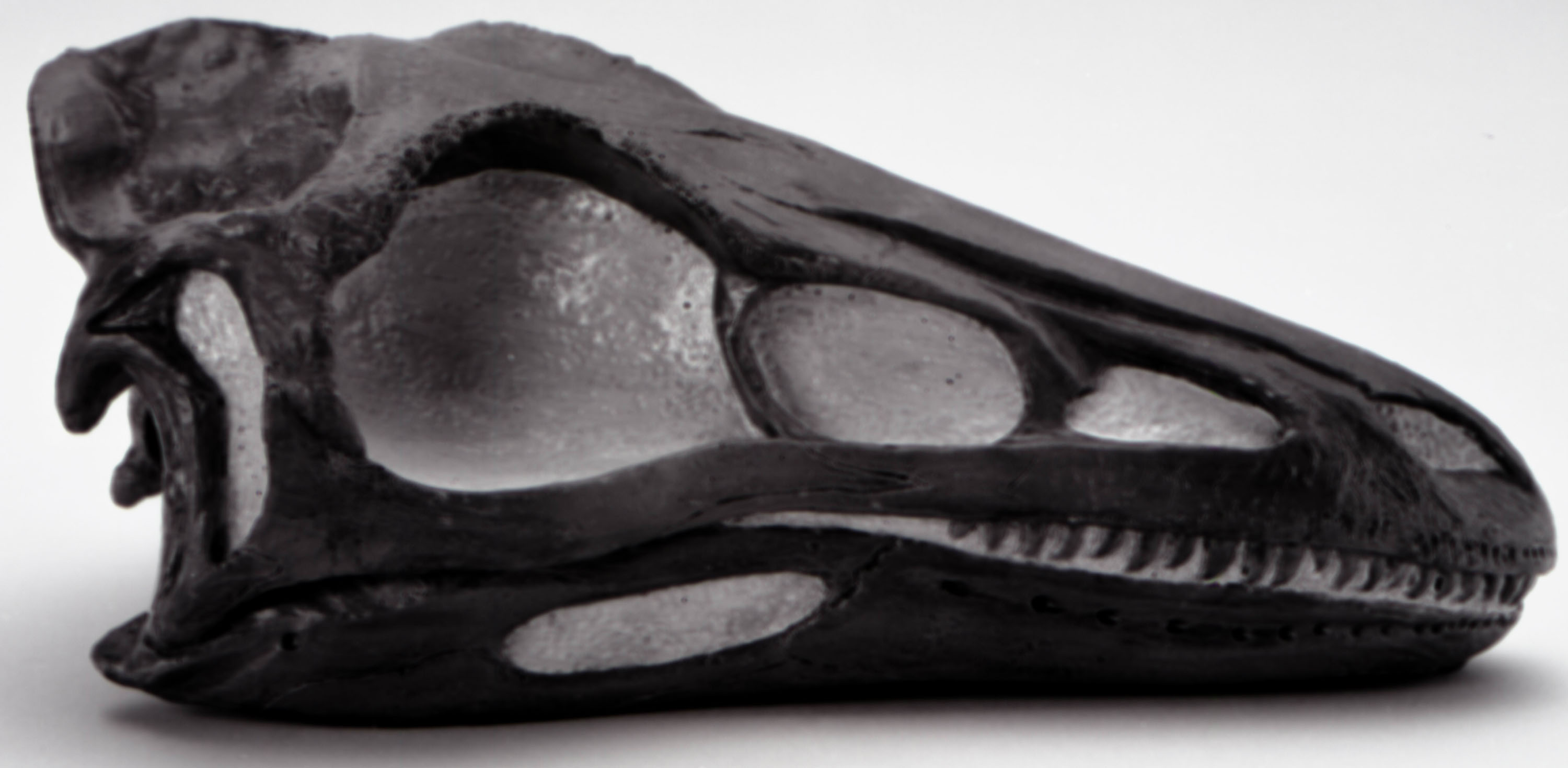

РИСУНОК 6. Реконструированный череп STENONYCHOSAUBUS INEQUALIS
(A - NMC neg. 81-7494,
вид сбоку; B - 81-7493, вид сверху).
Позвоночный столб представлен только фрагментом тела одного
предкрестцового (спинного) позвонка у NMC 12340, вместе с дистальным хвостовым
позвонком у этого образца и ещё одним у NMC 8539. В связи с этим его реконструкция
вызвала существенные затруднения наряду с вероятностью ошибки. Реконструированная
длина фрагмента тела позвонка в сравнении с формой позвонков, сохранившихся
у Saurornithoides mongoliensis (Russell 1969), Deinonychus antirrhopus
(Ostrom 1969B, рис. 34) и Saurornitholestes langstoni (Sues 1978),
составляет, по оценкам, 36 мм. Длина позвоночника до крестца рассчитана в соответствии
с размерами позвонков у AMNH 3015 (D. antirrhopus, Ostrom 1969B, таблица
4), с добавлением по 3 мм для каждого межпозвоночного диска. Общая длина тел
шести крестцовых позвонков оценена в соответствии с размерами тел позвонков
в крестце S. mongoliensis (Russell 1969, таблица 7). Проксимальные
хвостовые позвонки были реконструированы на основе таковых от S. junior
(Barsbold 1974B, вкл. 4, Рис. 2-3), также с привязкой по масштабу к ожидаемой
длине тел позвонков в основании хвоста у NMC 12340. Во всех случаях экстраполяции
приблизительно линейные, поскольку доступных материалов недостаточно, чтобы
устанавливать на их основе аллометрические тенденции. Сходство между длинами
тел позвонков задней части спины, крестца и основания хвоста у AMNH 6516 (S.
mongoliensis) интерпретируется в данном случае как свидетельство того,
что серия хвостовых позвонков вышеупомянутого образца S. junior включает
основание хвоста.
Из элементов передней конечности у NMC 12340 полностью сохранилась лишь локтевая
кость. Длина плечевой кости реконструирована в соответствии с размерами плечевой
и локтевой костей у D. antirrhopus (Ostrom 1969B, таблица 10). Фрагменты
элементов фаланг пальцев и когтевых фаланг позволяют предположить, что размер
кисти у NMC 12340 составлял примерно три четверти от таковой у YPM 5206 (D.
antirrhopus, Ostrom 1969B, рис. 62).
Таз был реконструирован на основе известного для Saurornithoides (Barsbold
1977, Рис. 1-7). Значения диаметра бедренной кости у NMC 12340 и AMNH 6516 (S.
mongoliensis) использовались для привязки по масштабу длины бедра и голени
+ таранной кости по оценке для последнего образца (Russell 1969, таблица 8)
к пропорциям реконструкции скелета. Ступня реконструирована по образцам NMC
8539 и NMC 12340. Размеры реконструированных частей скелета даны в таблице 2.
Мягкие ткани Stenonychosaurus inequalis были смоделированы непосредственно
на реконструкции скелета, что позволило осуществить проверку размеров, которая
в ином случае будет невозможна. Результаты этой проверки можно увидеть, если
сравнить модель в масштабе 1:7 (рис. 7), которая была изготовлена в процессе
отливки частей скелета, с законченной моделью в натуральную величину (рис. 8).
Наличие скелетного каркаса также повысило скорость, точность и достоверность,
необходимые для завершения реконструкции формы тела. Предварительная уменьшенная
модель требовалась главным образом для того, чтобы определить сбалансированную,
близкую к прижизненной жизни позу для реконструкции скелета.

РИСУНОК 7. Предварительная уменьшенная модель (1:7) формы тела
STENONYCHOSAURUS INEQUALIS,
вид сбоку (NMC neg. 81-1665).

РИСУНОК 8. Готовая полноразмерная модель (1:1) формы тела STENONYCHOSAURUS
INEQUALIS,
вид сбоку (NMC neg. 81-1244).
Мускулатура в целом соответствует описанной Ромером (Romer,
1956) для рептилий и изменена в целях соответствия строению теропод, а миология
тазовой области адаптирована на основе таковой у орнитомимид, реконструированной
Расселлом (Russell 1972). Примерные очертания мускульных масс были смоделированы
из пучков древесной шерсти, которые были покрыты папье-маше и дополнены тонким
слоем пластилина. Рёбра, связанные с образцом NMC 12340, определили форму тела
в поперечном сечении, а дистальные элементы таза определили очертания его брюшной
полости. Следует отметить, что тазовый канал у заврорнитоидов имеет такие же
относительные размеры, как у орнитомимид (см. также Russell 1969, стр. 608).
Для того, чтобы приблизительно смоделировать возможную форму трахеи, была использована
гибкая пластмассовая трубка. Фотографии больших ящериц (Schmidt and Inger 1957,
вкл. 33, 34, 52; рисунки на стр. 171, 172) дали нам детали, которые были использованы
в текстуре кожи, на кожных складках, веках и губах. Сочленения между костями
ступни у Stenonychosaurus, как и у многих других теропод, очень сильно
напоминают таковые у крупных наземных птиц; была близко воспроизведена внешняя
морфология ног бегающих птиц, в частности, казуара, (см. также Gilliard 1958,
вкл. 6, рис. на стр. 21; Stanek 1962, рис. на стр. 338-341). Очертания когтей
на кистях и ступнях были смоделированы по образцу отпечатков кератиновых чехлов,
сохранившихся у Archaeopteryx lithographica (Ostrom 1974, рис. 10;
Wellnhofer 1974, рис. 13) и Compsognathus longipes – его крошечного
современника из числа теропод (Ostrom 1978B, вкл. 9, рис. 5), но были немного
удлинены, чтобы лучше соответствовать их состоянию у современных крупных хищных
птиц (H. Ouellet, личное сообщение 1981). Внешний вид готовой модели показан
на рис. 8-12.
Реконструкция глаза заслуживает особого комментария. Приводимый Расселлом (Russell
1969, стр. 597) диаметр 52 мм предполагает, что глаз плотно прилегает к своду
полости глазницы. В настоящей реконструкции было взято более консервативное
значение диаметра 44 мм. Однако очевидно, что глаз у Stenonychosaurus
был крупный по сравнению с глазами современных наземных позвоночных, у которых
максимальным диаметром 50 мм обладает страус (Walls 1942, стр. 642). Средний
диаметр склеротических колец, указанных для различных таксонов динозавров, превышает
диаметр глаза страуса, и только у Psittacosaurus приблизительно достигает
размеров человеческого глаза (таблица 3). Почему динозавры обладали такими большими
глазами? Были ли они главным образом ночными существами, чтобы избегать тем
самым тепловой нагрузки, которую они в ином случае получали бы в случае дневной
активности? Были ли светопропускающие свойства атмосферы иными в те времена?
Случаю Stenonychosaurus с его сильно развитыми хищническими когтями
могут соответствовать следующие наблюдения (Walls 1942, стр. 209):
«Превосходная острота зрения дневного позвоночного часто позволяет ему сохранять значительную разницу между его вооружённостью и обороноспособностью его добычи – как в случаях, когда ястреб хватает подвязочную змею, или тиранн ловит муху. Ночной хищник должен обладать превосходящим оружием, поскольку обычно он должен бороться на более равных условиях с добычей значительно превосходящих его размеров».

РИСУНОК 9. Готовая модель тела STENONYCHOSAURUS INEQUALIS в
натуральную величину,
вид спереди и сбоку (NMC neg. 81-1243).

РИСУНОК 10. Модель головы STENONYCHOSAURUS INEQUALIS, вид сбоку
(NMC neg. 81-1242).
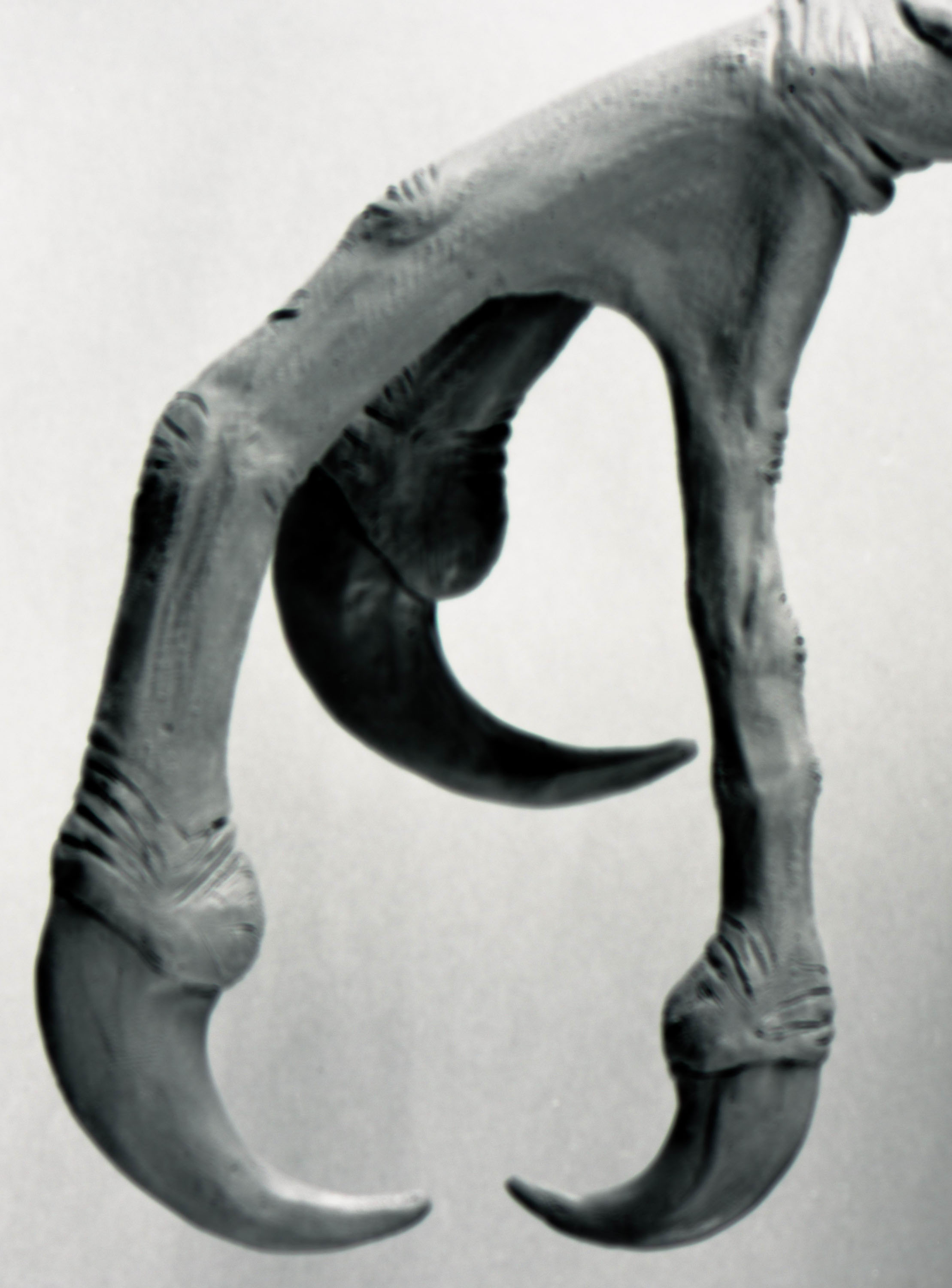
РИСУНОК 11. Модель кисти STENONYCHOSAURUS INEQUALIS, вид сбоку
(NMC neg. 81-1246).

РИСУНОК 12. Модель ступни STENONYCHOSAURUS INEQUALIS, вид с
медиальной стороны
(NMC neg. 81-1249).
Сходство между разбросом значений энцефализации для плотоядных
млекопитающих и травоядных, на которых они охотились (Radinsky 1978), позволяет
предположить, что мелкий хищный динозавр питался млекопитающими со сходным уровнем
развития мозга, которые, возможно, сами были более активными по ночам. Таким
образом можно постулировать, что Stenonychosaurus был сумеречным или
ночным хищником с необычайно острым зрением по сравнению с современными наземными
позвоночными. Гекконы известны остротой своего зрения (Bellairs 1970, стр. 350),
и в соответствии с этим глаз на реконструкции скопирован с глаза Gekko gecko
(Schmidt и Inger 1957, вкл. 22-23). Он был отлит из биопластика в виде полушария,
а радужная оболочка и зрачок были нарисованы на плоской внутренней поверхности.
С модели был сделан резиновый латексный слепок; он укреплён составной стекловолоконной
подкладкой, расположенной так, что на слепке остался лишь один узкий шов. Внутренние
поверхности слепка были покрыты стекловолокном, укреплённым эпоксидной смолой,
в которую для придания дополнительной жёсткости был вставлен металлический каркас.
В ступнях использовалась иная смесь эпоксидной смолы, укрепляющая контакт между
ступнями и основанием, который иначе был бы слабым. Отвердевший материал остался
достаточно гибким, чтобы позволить снять латексный слепок с кистей и ступней,
но достаточно прочным, чтобы слепок мог выдержать обращение с ним. Слепок был
смонтирован на основании, моделирующем испещрённые рябью поверхности, встречающиеся
в позднемеловой формации каньона Хорсшу в Альберте. Он был раскрашен вручную
и из распылителя масляными красками, чтобы его окраска напоминала узоры, типичные
для крупных рептилий, населяющих лесные местообитания (см. также Schmidt и Inger
1957), аналогичные тем, которые предположительно предпочитал Stenonychosaurus
(Béland and Russell 1978). Следует особо отметить, что не делалось никаких попыток
придать окрашенному слепку (рис. 13) облик сообразительного или интеллектуально
развитого существа. Размеры готовой реконструкции приводятся в таблице 4. Для
завершения проекта потребовалось семь месяцев.
Поскольку Stenonychosaurus фигурировал в дискуссиях о параллельном приобретении разума пресмыкающимися, при помощи готовой реконструкции было заново оценено соотношение веса его мозга и тела. Было установлено, что модель вытесняла 46,6 кг воды. Если исходить из того, что плотность тела ныне живущего животного равна 0,8 или 0,9 (Colbert 1962, стр. 5), это означает вес 37,3 или 41,9 кг, что оказывается меньше, чем 45,3 кг по оценке Расселла (Russell 1969, стр. 599) на основании пропорций бедренной кости. Вес мозга оценивался в 37 (Hopson,

РИСУНОК 13. Раскрашенный слепок реконструированной формы тела
STENONYCHOSAURUS INEQUALIS,
вид сбоку (NMC neg. 81-3251).
1980, таблица 1, стр. 297) и 45 граммов (Russell 1969, стр. 599). Если использовать приводимое Джерисоном (Jerison 1973) общее соотношение между весом мозга (E, граммы) и весом тела (P, граммы) у млекопитающих:
| E ожидаемый = 0,12 P 2/3 | (уравнение 1) |
и определения коэффициента энцефализации (EQ) как
| EQ = E наблюдаемый/E ожидаемый | (уравнение 2), |
то для Stenonychosaurus были получены значения энцефализации,
колеблющиеся между 0,24 и 0,34 (таблица 5). Это близко к разбросу значений энцефализации,
наблюдаемому среди цесарок и дроф, среди броненосцев и опоссумов (Crile и Quiring
1940), или среди примитивных насекомоядных (Jerison 1973, таблица 10.3).
Хотя относительный размер мозга у Stenonychosaurus был гораздо больше,
чем у большинства динозавров (Hopson 1980), другие мелкие тероподы также достигли
сопоставимых уровней энцефализации (см. также Russell 1972, Hopson 1977, Sues
1978). Поскольку эти существа и мелкие динозавры в целом так плохо известны,
вполне вероятно, что формы, которые ещё только предстоит описать, потеснят Stenonychosaurus
с места обладателя наивысшей степени развития головного мозга, зарегистрированной
для динозавров. Если представленная здесь реконструкция (рис. 12) приблизительно
верная, это было вне всяких сомнений интересное животное. При весе около 40
кг это был один из самых мелких динозавров, известных с запада Канады. Его крупные
глаза предполагают подвижный образ жизни (Walls 1942, стр. 173), об этом же
свидетельствуют стереоскопическое зрение, длинные ноги и лёгкое телосложение.
Хвост был самым гибким в основании; это подразумевает, что он контролировал
вращательный момент тела путём его поднятия или вытягивания на быстрых поворотах.
Форма локтевой кости указывает, что предплечье могло вращаться, а строение запястья
предполагает, что третий палец, возможно, мог противопоставляться по отношению
к двум другим (Russell 1969). Плотоядные повадки явственно выражены в строении
конечностей, где кисти были приспособлены для удержания жертвы, пока ей вырывали
внутренности когти на внутренней стороне ступни.
Stenonychosaurus был в высшей степени прогрессивным животным для своего
времени, и, возможно, он не показался бы слишком неуместным среди австралийской
фауны наших дней.
Для последних 600 миллионов лет эпохи фанерозоя можно составить
график, представляющий максимальный уровень энцефализации, известный для организмов,
живущих на нашей планете (Russell 1981, рис. 1). Человек и его непосредственные
предшественники попадают на этот график, а Stenonychosaurus сильно
приблизился к нему во времена позднего мезозоя. S. inequalis (NMC 12340)
жил примерно за 12 миллионов лет до конца эры рептилий. Хотя отдельные кости
заврорнитоидов известны из самых молодых мезозойских отложений (K. Carpenter,
личное сообщение, 1981), не собрано никаких скелетных фрагментов, которые были
бы достаточно полными для получения оценки соотношения веса мозга и тела. Шестьдесят
четыре миллиона лет назад динозавры, в том числе родословная ветвь заврорнитоидов,
исчезли с лица нашей планеты во время катастрофы, исследование масштабов которой
началось лишь в настоящее время. Было бы весьма любопытно узнать, как признаки
заврорнитоидов – крупный размер мозга, стереоскопическое зрение, противопоставляющиеся
пальцы и двуногое положение тела – менялись на протяжении оставшегося времени
мезозойской эры, если это имело место. Также было бы интересно поразмышлять
с позиций качественной оценки того, как могли бы выглядеть потомки S. inequalis,
если бы они пережили вымирание конца мелового периода и приобрели коэффициент
энцефализации, как у Homo sapiens (около 7,5; см. также Jerison 1973,
таблица 16.3), двигаясь в своё будущее на протяжении 76 миллионов лет по кривой
максимальной энцефализации.
Полушария мозга S. inequalis были примерно треугольных очертаний при
взгляде сверху, больше напоминая таковые у птиц, чем более округлые очертания,
наблюдаемые у антропоидов (см. также Russell 1969, рис. 3; Romer 1962, рис.
388; Gregory 1951, рис. 23.64). Эти общие очертания сохранялись при увеличении
объёма эндокрана до 1100 мл, что равняется таковому у черепа маленького женского
скелета из коллекции отдела палеобиологии. По мере увеличения объёма эндокрана
относительные размеры лицевой области уменьшаются, и в качестве ориентира для
создания модели гипотетического черепа были выбраны краниофациальные размеры
куриного эмбриона (Goodrich 1958, рис. 622). Зубы у заврорнитоидов редуцированы
по отношению к зубам дромеозавров, и было принято решение убрать их вообще,
заменив ороговевшими окклюзионными поверхностями, как у черепах и в меньшей
степени у орнитомимид. Такие педоморфные особенности, как относительно крупный
мозг и редуцированная зубная система, также характерны для высших антропоидов
(Gould 1977). Повышенные требования к обмену веществ со стороны крупного мозга
(Sokoloff 1976) согласуются с приобретением эндотермии и потребностью в богатой
энергией пище. Потенциальной опасности гниения зубов, если предположить наряду
с плотоядностью поедание фруктов, удаётся избежать путём устранения зубов. Наружные
ноздри в модели был сдвинуты вверх, чтобы дать место вторичному нёбу – известно,
что у орнитомимид случилось то же самое (Russell 1972, стр. 399). Высокий уровень
энцефализации по определению подразумевает, что голова будет тяжёлой по отношению
к общему весу тела и породит особые проблемы её поддерживания. У Pachycephalosaurus
сравнительно массивный, хотя и состоящий из кости череп уравновешен на затылочном
мыщелке и большом затылочном отверстии, которые сместились к центру нижней поверхности
черепа (Brown and Schlaikjer 1943, вкл. 39, рис. 1). Вышеупомянутые признаки
были объединены в модели гипотетического черепа (рис. 14), на который наложили
рисунок швов между частями черепа S. inequalis. Размеры орбит были
сохранены из уважения к крупному размеру глаз у динозавров в целом. Боковые
височные окна S. inequalis были закрыты отростками чешуйчатой, посторбитальной
и скуловой костей, а квадратно-скуловая кость исчезла. На верхней челюсти сохранилась
лишь единственная пара предглазничных окон, а наружные нижнечелюстные окна исчезли.
Остеологические упрощения подобного рода часто наблюдаются в группах позвоночных,
которые продолжали существовать на протяжении многих десятков миллионов лет.
В соответствии с типичным рептильным состоянием нижнечелюстной сустав располагается
ниже окклюзионной поверхности, а не выше её, как у большинства млекопитающих.
Архитектура челюстей развита не слабее, чем у нашего собственного вида.
Положение позвоночного столба у гипотетического потомка S. inequalis с
высокоразвитым мозгом более проблематично. Возможно, увеличенную голову можно
успешно поддерживать путём перераспределения веса тела в положение за ногами
и формирования состоящей из связок подвески вдоль верхней стороны груди и шеи
(это приспособление наглядно заметно у бескилевых птиц и зауропод). Однако опыт
множества людей по всему миру говорит о том, что большие грузы значительно легче
носить, уравновесив их на голове, чем повесив на спину или держа в руках. Если
использовать этот пример как аналогию при выявлении ограничивающего условия
для опоры черепа, кажется разумным предположить, что энергетически эффективнее
уравновесить увеличенную голову на вертикальной шее, чем подвесить её на конце
горизонтальной шеи при помощи мускулов и связок. Браун и Шлейкер (Brown and
Schlaikjer 1943, стр. 138) утверждали, что Pachycephalosaurus был габитуально
двуногим, и делают вывод о том, что он обладал более выпрямленной позой, чем
другие двуногие птицетазовые динозавры. К сожалению, посткраниальный скелет
этого динозавра неизвестен. Тенденция к более прямой постановке головы на вершине
позвоночного столба наблюдается в ряду антропоидов с увеличивающимся размером
мозга (Adams and Moore 1975). Укороченная шея уменьшила бы перегрузки, испытываемые
чувствительным к толчкам мозгом, которые будут следствием движений шеи и тела
(см. также Oldendorf and Zabielski 1981).
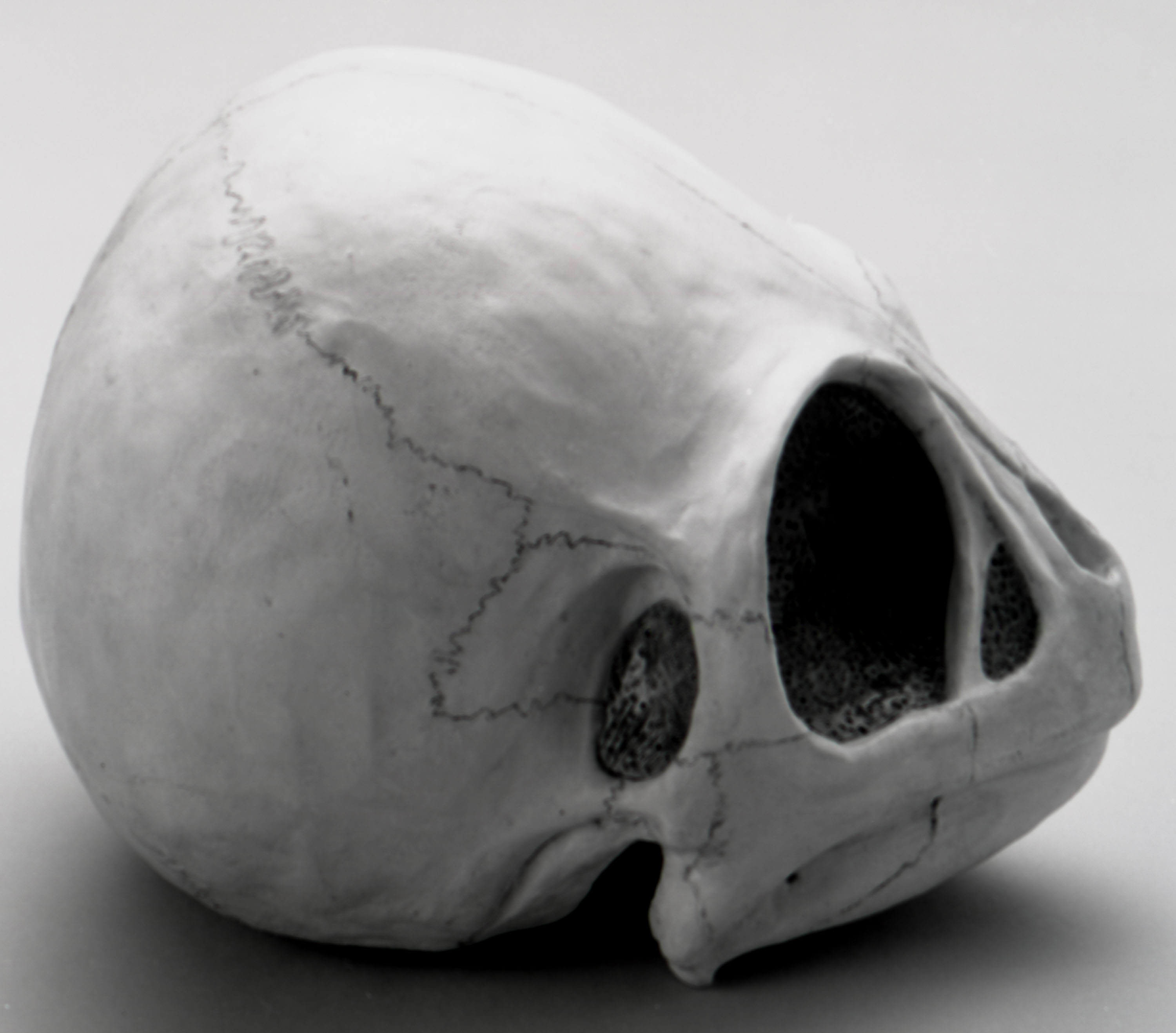
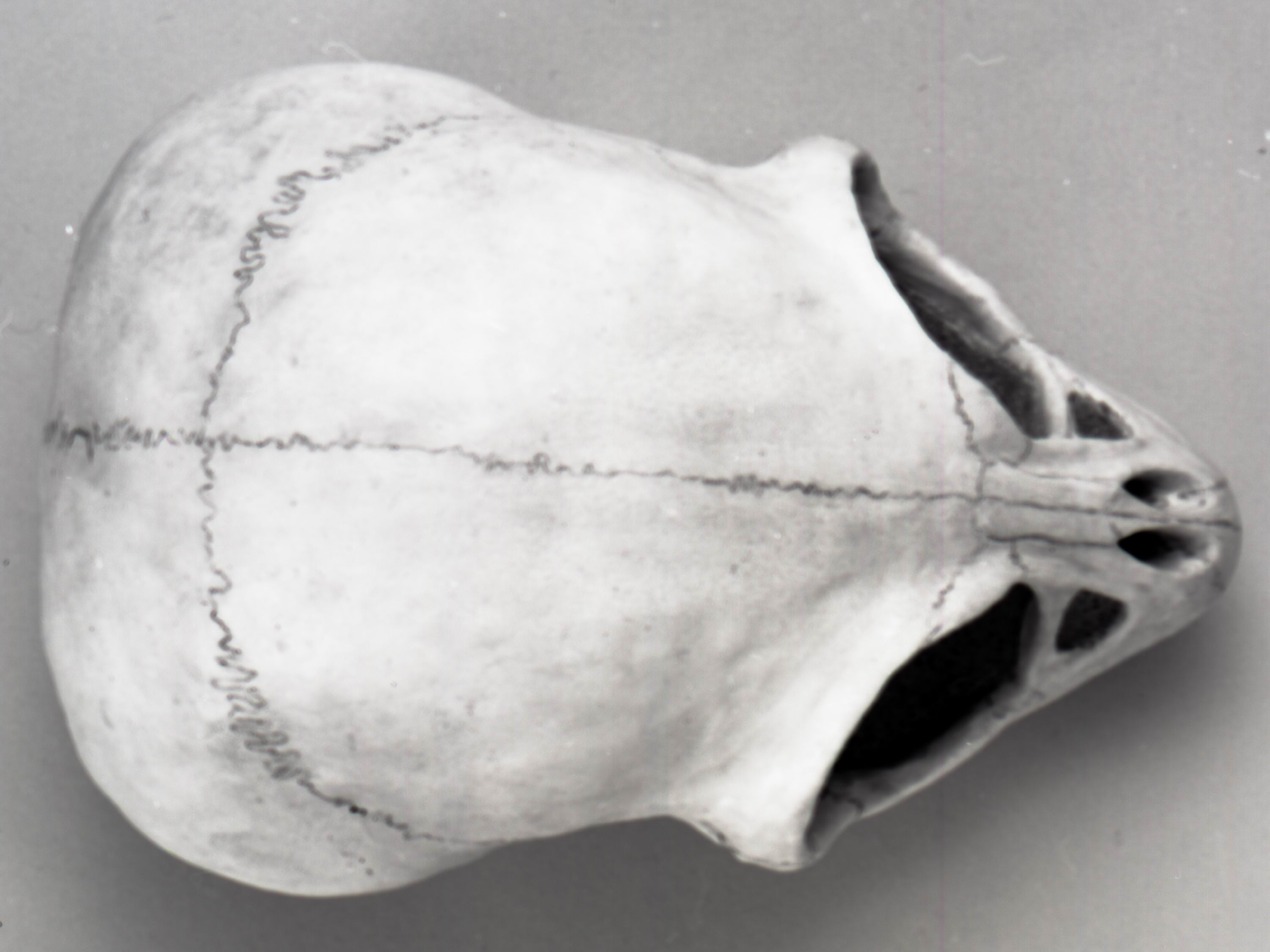
РИСУНОК 14. Модель черепа динозавроида (А – NMC neg. 81-7491,
вид сбоку;
B – 81-7490, вид сверху).
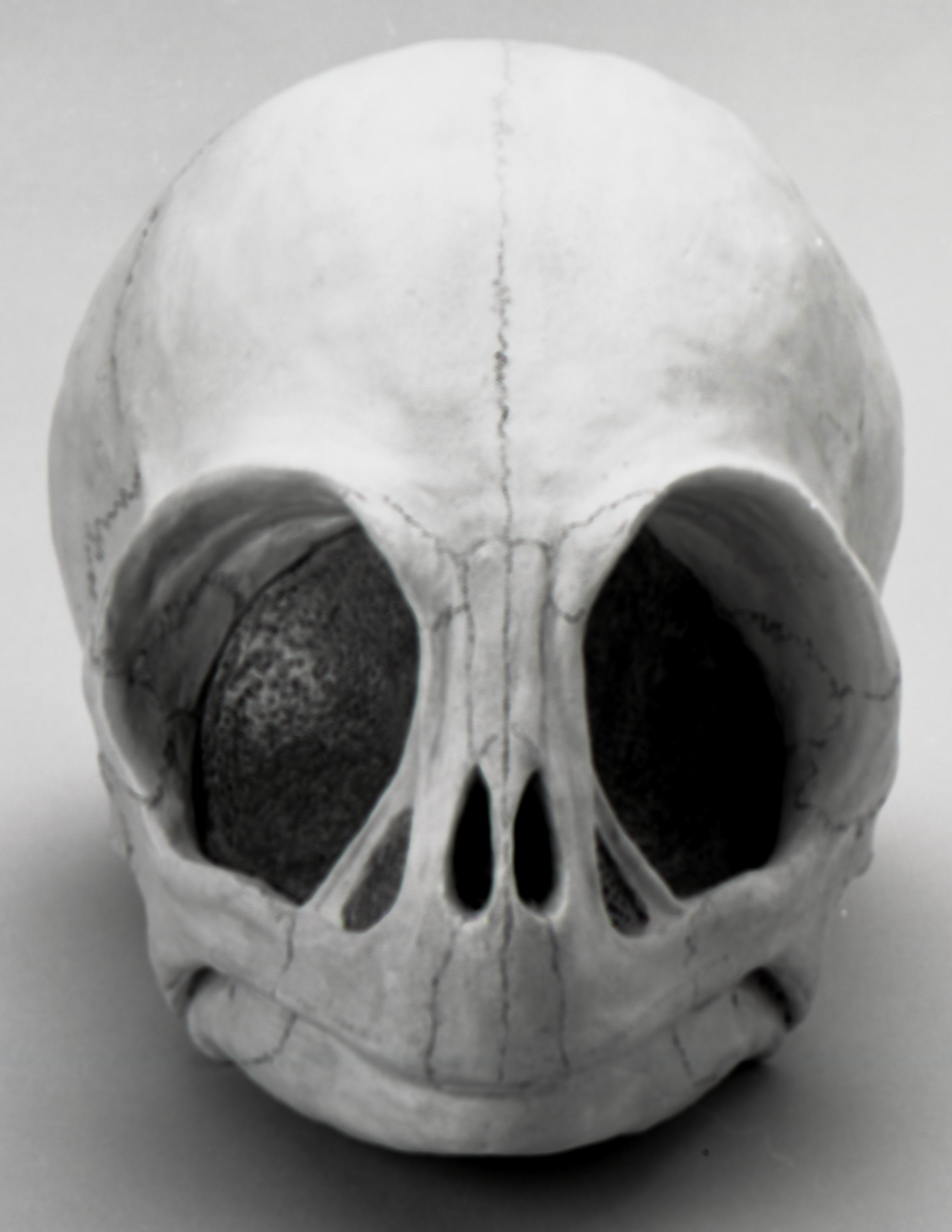
РИСУНОК 14. (C - 81-7489, вид спереди).
Если говорить об отделах позвоночного столба ниже шеи, то известно,
что приобретение вертикального положения тела предшествовало увеличению мозга
у гоминид в процессе филогенеза (White 1980). Возможно, при уровне энцефализации,
близком или чуть большем, чем у человекообразных обезьян, способность использовать
инструменты становится важнейшим фактором в процессе отбора (Washburn 1978,
Lewin 1981). Энергетически эффективный способ передвижения (Rodman and McHenry
1980; затраты на перемещение единицы веса у человека низкие по отношению к таковым
у многих наземных позвоночных) также может быть выгодным при поиске мест изобилия
богатой энергией пищи (собирательство-охота), неравномерно распределённых по
неоднородным экосистемам. Если эти предположения верны, то затраты энергии,
необходимые для поддержания несколько увеличенной головы и передних конечностей
в рамках строения скелета Stenonychosaurus стали бы больше, а позвоночный
столб, ориентированный в передне-заднем направлении, увеличил бы угловую инерцию
разворачивающегося тела, ухудшая тем самым способность бросать метательные снаряды.
Преодолеть эти трудности наилучшим образом можно, развернув позвоночный столб
вверх, а не сдвинув центр тяжести назад. Тогда ноги, туловище и руки можно задействовать
в качестве системы рычагов в процессе приложения силы к инструментам (см. также
Washburn and Moore 1980, стр. 69-79).
Предварительная скелетная модель для гипотетического потомка S. inequalis,
или «динозавроида», была создана на основе модели черепа и вертикального позвоночного
столба с укороченной шейной областью. У заврорнитоидов (Barsbold 1977, Рис.
1-7) длинная ось подвздошной кости направлена под прямым углом по отношению
к таковым у вентральных тазовых элементов, и её передняя лопасть расширена не
настолько, чтобы выходить за рамки состояний, типичных для теропод, живших одновременно
с ними. У людей передняя лопасть подвздошной кости сильно расширена, а ветви
лобковой и седалищной костей развёрнуты постериорно (в морфологическом смысле)
относительно положения соответствующих структур у заврорнитоидов. У Segnosaurus,
особого теропода из мела Монголии, строение таза примерно промежуточное между
этими двумя крайностями (см. также Perle 1979, рис. 5) – возможно, как результат
близкой к вертикальной ориентации позвоночного столба у этого рода. Тазовый
канал гипотетического динозавроида был бы расширен, чтобы облегчить рождение
молодняка с высокоразвитым мозгом. Было бы вполне возможно вывести таз, похожий
на гоминидный, из тазовой морфологии заврорнитоидов, и такой таз, соответственно,
прикреплялся бы к заднему концу предкрестцового участка позвоночного столба
скелета модели. Осевой скелет заканчивается видимым снаружи рудиментом хвоста.
Stenonychosaurus был специализированным бегающим животным, и изменения
в строении ноги, которые необходимы вертикально расположенному позвоночнику,
вероятно, ухудшат его способность бегать (см. также Lovejoy 1981). Возможно,
это ограничение было бы компенсировано селективными преимуществами, которые
являются следствием более высокоразвитой центральной нервной системы. Неустойчивость,
порождаемая положением колена и интертарзального сустава над субстратом, может
оказаться энергетически дорогой для медленно передвигающихся существ из-за необходимости
значительной «тонкой настройки» мускулатуры. Поэтому плюсна модели была укорочена
и поставлена в горизонтальном положении, а пяточная область была удлинена (см.
также Lessertisseur and Jouffroy 1975). У заврорнитоидов есть лишь два элемента
предплюсны (Barsbold 1974B, рис. 5); возможно, пятая метатарзалия (Russell 1969,
рис. 13) могла бы превратиться в обызвествлённое окостеневшее образование. Интертарзальный
сустав у модели вогнутый, он такой же у заврорнитоидов и бескилевых птиц. Пропорции
ноги – как у H. sapiens, если исходить из предположения, что у нашего
вида нога хорошо подходит для прямохождения. Кнемиальный гребень большеберцовой
кости в верхней части передней поверхности голени, по строению как у теропод,
заменяет коленную чашечку в качестве места прикрепления мышц-абдукторов голени.
Не-хищнические пальцы ног модели выполнены по образцу пальцев задних лап древесного
кенгуру (Dendrolagus, см. также Gregory 1951, рис. 18.37Z), которые
в иных отношениях в целом напоминают пальцы S. inequalis. Умелое использование
орудий труда сделало бы излишней необходимость в оружии телесной природы – так
же, как нашими собственными предшественниками были утрачены увеличенные клыки.
У гоминид плечи такие же широкие или шире, чем у способных к брахиации антропоидов
(см. также Gregory 1951, Рис. 24.1, 24.2); возможно, это следствие хорошо скоординированного
умения бросать метательные снаряды (Brues 1959). На скелете модели были сделаны
хорошо выраженные плечи, которые прилегают к грудине антеромедиально при помощи
широких, но удлинённых коракоидов, а не ключиц. Грудная клетка узкая и выдающаяся
– такая же, как у S. inequalis. Соотношение длин плеча и предплечья
похоже на наблюдаемое у орнитомимид (Osborn 1917, рис. 8), а также у гоминид.
Кисть трёхпалая, как у S. inequalis, и третий палец находится в положении большого
пальца. Пальцы заканчиваются уплощёнными когтевыми фалангами.
После этого скелетный каркас динозавроида был «восстановлен» и с него была сделана
отливка так же, как у Stenonychosaurus (Рис. 15-19). Рост модели в
выбранной позе в стоячем положении составляет 1350 мм. Было установлено, что
форму для отливки можно было целиком заполнить 31,7 литрами песка. Если исходить
из предположений, что мозг заполнял 80% полости эндокрана, а удельный вес тела
составлял 0,9, это означает живой вес 28,5 кг и коэффициент энцефализации 7,1
(уравнения 1 и 2). Таким образом, уровень энцефализации динозавроида лежит в
пределах, указанных для человека (Jerison 1973, таблица 16.3). Хотя значения
веса тела, указанные по двум моделям, не слишком отличаются (28 против 40 кг),
это указывает на то, что высокий уровень энцефализации был достигнут посредством
как утраты тканей тела, так и увеличения веса мозга. Чтобы закончить вторую
модель, потребовалось три с половиной месяца. Наличие или отсутствие некоторых
образований в анатомии его мягких тканей требует дополнительного комментария.

РИСУНОК 15. Модель головы динозавроида, вид спереди и сбоку (NMC neg. 81-7485).

РИСУНОК 16. Модель кисти динозавроида, вид с медиальной стороны (EMC neg. 81-7488).

РИСУНОК 17. Модель ступни динозавроида, вид с медиальной стороны (NMC neg. 81-7487).

РИСУНОК 18. Готовые модели STENONYCHOSAURUS INEQUALIS и динозавроида
(A - NMC neg. 81-7483, вид с правого бока).

РИСУНОК 18. (B - 81-7481, вид спереди).

РИСУНОК 18. (C - 81-7482, вид сзади).

РИСУНОК 19. Готовая модель динозавроида, вид с правого бока (NMC neg. 81-8401).
Текстура кожи и общая окраска были взяты с изменениями с реконструкции
Stenonychosaurus. На голове глаза были развёрнуты антеромедиально как
следствие расширения черепной коробки, результатом чего стало более значительное
перекрывание полей зрения. Вертикальный щелевидный зрачок был заимствован у
предыдущей модели, но его фестончатые края, типичные для некоторых гекконов,
были убраны. Серьга, или кожная складка под нижней челюстью – вторичный половой
признак. Наружная ушная раковина отсутствует, что типично для рептилий и птиц.
Точно так же на брюшной стороне тела нет сосков; можно предположить, что молодняк
поедал пищу, отрыгиваемую родителями. Широкий тазовый канал - препятствие для
успешного двуногого хождения (Lovejoy 1975), и его диаметр у человека сведён
к минимуму благодаря рождению молодых особей в относительно незрелом состоянии,
когда голова меньшего размера (Passingham 1975, p. 81). Наличие тканей яйца
лишь усугубило бы эту проблему. Поскольку у некоторых рептилий (в частности,
у некоторых сцинков, см. также Bellairs 1970, стр. 449-452) имеется хорошо развитая
плацента, на животе динозавроида был сделан пупок. Похожая на ягодицы масса
мускулатуры, покрывающая основание таза – это следствие редукции хвоста и замещения
лежащих в глубине хвостовобедренных мышц подвздошно-бедренными, начинающимися
на расширенном переднем крыле подвздошной кости (см. также Russell 1972, рис.
7-8; Sigmon 1975, рис. 3-4). Отсутствуют видимые снаружи половые органы, которые
у большинства рептилий и птиц обычно находятся внутри клоаки.
Реконструкцию Stenonychosaurus можно подвергнуть проверке путём обнаружения
более полного материала. В случае динозавроида это неприменимо, возможно, даже
отчасти, поскольку темп изменения максимального значения уровня энцефализации
в позднемеловую эпоху мог быть недостаточно быстрым (см. также Russell 1981),
чтобы продемонстрировать его обнаружимое увеличение у потомков S. inequalis
до завершения эпохи. Форма тела Homo sapiens возможна, потому что она
существует. В той степени, в какой она приближается к таковой у динозавроида,
последний также становится вероятной биофизической конфигурацией. Но не лучше
ли будет объяснить это сходство результатом «ортогенетической» тенденции? Возможно.
Однако, если это так, следует также допустить, что в пределах спектра вариантов
морфологии, представленных среди последних динозавров мелового периода, существовала
мозаика признаков, которые появились параллельно многим особенностям, наблюдаемым
у млекопитающих и у филогенетических предшественников человека. Этим обстоятельствам
следует давать оценку в свете знания о повсеместности явления параллельной эволюции
в истории жизни на Земле. С другой стороны, мог ли динозавроид действительно
быть слишком «рептильным», чтобы представлять собой приемлемую схему строения?
Разве глаза не должны быть меньше, а зрачки – более округлыми? Разве не были
бы внешние уши полезными для концентрирования звука на барабанной перепонке?
Не слишком ли сильны челюсти, и не длинновата ли морда? Совместима ли сильно
выдающаяся узкая грудь с наличием плеч?
Динозавроид может представлять собой решение в ответ на физические и физиологические
стрессы, которые накладывает на организм позвоночного сильно гипертрофированный
мозг в наземной среде обитания. Если это верно, то его форма в целом может иметь
биологическое значение, аналогичное, например, птицеподобной форме птиц, летучих
мышей и птерозавров, рыбоподобной форме костистых рыб, акул и морских свиней,
или «лошадиной» форме лошадей и некоторых вымерших южноамериканских копытных
(литоптерн). Наличие этой формы тела у Homo sapiens показывает, что
решение существует. Однако оно может быть не единственным. Мы приглашаем наших
коллег к выявлению альтернативных решений.
|
РОД |
МЕСТО НАХОДКИ |
ТИПОВОЙ МАТЕРИАЛ |
АВТОР(Ы) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Длина черепа (от предчелюстной кости до затылочного мыщелка) | 242 мм |
| Максимальная ширина черепа | 145 |
| Длина шейного отдела (10 позвонков) | 405 |
| Длина спинного отдела (12 позвонков) | 465 |
| Длина крестцового отдела (6 позвонков) | 222 |
| Длина хвостового отдела | 1680 |
| Длина плечевой кости | 157 |
| Длина локтевой кости | 130 |
| Длина II пальца (кисть) | 192 |
| Длина подвздошной кости | 268 |
| Высота таза и крестца | 370 |
| Длина бедра | 330 |
| Длина голени + таранной кости | 405 |
* Эти измерения являются основанием для данной реконструкции скелета, и ещё могут облегчить поиск ошибок, когда будет доступен более полный материал.
ТАКСОН |
ПРИБЛИЗИТЕЛЬНЫЙ ДИАМЕТР |
ИСТОЧНИК |
| Theropoda | ||
|
|
|
|
|
|
| Sauropoda | ||
|
|
|
|
|
|
|
|
|
| Ornithopoda | ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ceratopsia |
||
|
|
|
|
|
|
|
|
|
| СРЕДНЕЕ ЗНАЧЕНИЕ |
|
| Длина (от морды до анального отверстия) | 1460 мм |
| Длина (от анального отверстия до кончика хвоста) | 1620 |
| Общая длина (в данной позе) | 2805 |
| Высота грудной клетки | 384 |
| Ширина грудной клетки | 240 |
| Высота в крестце | 980 |
| Высота на уровне головы | 1190 |
| ВЕС МОЗГА (ГРАММЫ) | ВЕС ТЕЛА (ГРАММЫ) | КОЭФФИЦИЕНТ ЭНЦЕФАЛИЗАЦИИ |
37 |
45 000 | 0,2439 |
| 37 | 41 940 | 0,2556 |
| 37 | 37 280 | 0,2767 |
| 45 | 45 300 | 0,2953 |
| 45 | 41 940 | 0,3109 |
| 45 | 37 280 | 0,3363 |
Дальнейшие пояснения см. в тексте на стр. 21
Adams, L.M., and W.J. Moore. 1975. Biomechanical appraisal of some skeletal
features associated with head balance and posture in the Hominoidea. Acta Anatomica
92: 580-594.
Barsbold, R. 1974A. Duel of the dinosaurs (in Russian). Priroda 1974(2): 81-83.
-- 1974B. Saurornithoididae, a new family of small theropod dinosaurs from Central
Asia and North America. Palaeontologia Polonica 30: 5-22.
-- 1976. On the evolution and systematics of late Mesozoic carnivorous dinosaurs
(in Russian). Transactions of the Soviet-Mongolian Paleontological Expedition
3: 68-75.
-- 1977. On the evolution of carnivorous dinosaurs (in Russian). Transactions
of the Soviet-Mongolian Paleontological Expedition 4: 48-56.
Barsbold, R., and A. Perle. 1979. Modification of the pelvis in the Saurischia
and parallel evolution in carnivorous dinosaurs (in Russian). Transactions of
the Soviet-Mongolian Paleontological Expedition 8: 39-44.
Béland, P., and D.A. Russell. 1978. Paleoecology of Dinosaur Provincial Park
(Cretaceous), Alberta, interpreted from the distribution of articulated vertebrate
remains. Canadian Journal of Earth Sciences 15: 1012-1024.
Bellairs, A. 1970. The life of reptiles. Universe Books, New York. 589 pp.
Bonaparte, J.F., and J.E. Powell. 1980. A continental assemblage of tetrapods
from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda
– Coelurosauria – Carnosauria – Aves) . Mémoires de la Société géologique de
France (n.s.) 139: 19-28.
Brown, B. 1917. A complete skeleton of the horned dinosaur Monoclonius
and description of a second skeleton showing skin impressions. American Museum
of Natural History Bulletin 37: 281-306.
Brown, B., and E.M. Schlaikjer. 1940. The structure and relationships of Protoceratops.
Annals of the New York Academy of Science 40: 133-266.
-- 1943. A study of the troodont dinosaurs with the description of a new genus
and four new species. American Museum of Natural History Bulletin 82: 115-150.
Brues, A. 1959. The spearman and the archer – an essay on selection in body
build. American Anthropologist 61: 457-469.
Colbert, E.H. 1962. The weights of dinosaurs. American Museum of Natural History
Novitates 2076: 1-16.
Colbert, E.H., and D.A. Russell. 1969. The small Cretaceous dinosaur Dromaeosaurus.
American Museum of Natural History Novitates 10024: 1-49.
Crile, G. , and D.P. Quiring. 1940. A record of the body weight and certain
organ and gland weights of 3690 animals. Ohio Journal of Science 40: 219-259.
Galton, P.M. 1973. Redescription of the skull and mandible of Parksosaurus
from the late Cretaceous with comments on the family Hypsilophodontidae. Royal
Ontario Museum Life Science Contribution 89: 1-21.
-- 1980. Dryosaurus and Camptosaurus, intercontinental genera
of upper Jurassic ornithopod dinosaurs. Mémoires de la Société géologique de
France (n.s.) 139: 103-108.
Gilliard, E.T. 1958. Living birds of the world. Doubleday, Garden City. 400
pp.
Goodrich, E.S. 1958. Studies on the structure and development of vertebrates.
Dover Publications, New York. 837 pp.
Gould, S.J. 1977. Ontogeny and phylogeny. Harvard University Press, Cambridge.
501 pp.
Gregory, W.K. 1951. Evolution emerging. MacMillan, New York. 1749 pp.
Hanzak, J. 1965. The pictorial encyclopedia of birds. Hamlyn, London. 582 pp.
(Русское издание: Ганзак, Ян «Иллюстрированная энциклопедия птиц», Прага, «Артия»,
1986)
Holland, W.J. 1924. The skull of Diplodocus. Memoirs of the Carnegie
Museum 9: 379-403.
Hopson, J. 1977. Relative brain size and behavior in archosaurian reptiles.
Annual Review of Ecology and Systematics 8: 429-448.
-- 1980. Relative brain size in dinosaurs. American Association for the Advancement
of Science Selected Symposium 28: 287-310.
Janensch, W. 1935. Die Schàdel der Sauropoden Brachiosaurus , Barosaurus
und Dicraeosaurus aus den Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica
Supplement 7, Reihe 1, Teil 2: 145-298.
Jerison, H.J. 1973. Evolution of the brain and intelligence. Academic Press,
New York. 482 pp.
Johanson, D.C., and M.A. Edey. 1981. Lucy, the beginnings of humankind. Simon
and Schuster, New York. 409 pp. (Русское издание: Джохансон Д., Иди М. «Люси.
Истоки рода человеческого» М., «Мир», 1984)
Lessertisseur , J., and F.K. Jouffroy. 1975. Comparative osteometry of the foot
of man and facultatively bipedal primates. In: Primate Functional Morphology
and Evolution. Edited by; R.H. Tuttle. Mouton Publishers, The Hague. pp. 327-340.
Lewin, R. 1981. Ethiopian stone tools are world's oldest. Science 211: 806-807.
Lovejoy, C. 1975. Biomechanical perspectives on the lower limb of early hominids.
In: Primate Functional Morphology and Evolution. Edited by: R.H. Tuttle. Mouton
Publishers, The Hague. pp. 291-326.
-- 1981. The origin of man. Science 211: 341-350.
Matthew, W.D. , and B. Brown. 1922. The family Deinodontidae, with notice of
a new genus from the Cretaceous of Alberta. American Museum of Natural History
Bulletin 46: 367-385.
Nowinski, A. 1971. Nemegtosaurus mongoliensis (Sauropoda) from the
uppermost Cretaceous of Mongolia. Palaeontologia Polonica 25: 57-81.
Oldendorf, W., and W. Zabielski. 1981. The buoyant brain. Science Digest 89:
94-96, 117.
Osborn, H.F. 1917. Skeletal adaptations of Ornitholestes, Struthiomimus,
Tyrannosaurus. American Museum of Natural History Bulletin 35: 733-771.
-- 1924A. Psittacosaurus and Protiguanodon: two lower Cretaceous
iguanodonts from Mongolia. American Museum of Natural History Novitates 127:
1-16.
-- 1924B. Three new Theropoda, Protoceratops zone, central Mongolia. American
Museum of Natural History Novitates 144: 1-12.
Ostrom, J.H. 1961. Cranial morphology of the hadrosaurian dinosaurs of North
America. American Museum of Natural History Bulletin 122: 33-186.
-- 1969A. A new theropod dinosaur from the lower Cretaceous of Montana. Yale
Peabody Museum Postilla 128: 1-17.
-- 1969B. Osteology of Deinonychus antirrhopus, an unusual theropod
from the lower Cretaceous of Montana. Peabody Museum of Natural History Yale
University Bulletin 30: 1-165.
-- 1974. Archaeopteryx and the origin of flight. Quarterly Review of
Biology 49: 27-47.
-- 1976. On a new specimen of the lower Cretaceous theropod dinosaur Deinonychus
antirrhopus. Museum of Comparative Zoology Breviora 439: 1-21.
-- 1978A. A new look at dinosaurs. National Geographic Magazine 154; 152-185.
-- 1978B. The osteology of Compsognathus longiceps Wagner. Zitteliana
4: 73-118.
Parks, W.A. 1928. Struthiomimus samueli a new species of Ornithomimidae
from the Belly River Formation of Alberta. University of Toronto Studies, Geological
Series 26: 1-24.
Passingham, R.E. 1975. Changes in the size and organization of the brain in
man and his ancestors. Brain, Behaviour and Evolution 11: 73-90.
Perle, A. 1979. Segnosauridae – a new family of theropods from the late Cretaceous
of Mongolia (in Russian). Transactions of the Soviet-Mongolian Paleontological
Expedition 8: 45-55.
Radinsky, L. 1978. Evolution of brain size in carnivores and ungulates. American
Naturalist 112: 815-831.
Rodman, P.S., and H.M. McHenry. 1980. Bioenergetics and the origin of hominid
bipedalism. American Journal of Physical Anthropology 52: 103-106.
Romer, A.S. 1956. Osteology of the reptiles. University of Chicago Press, Chicago.
772 pp.
-- 1962. The vertebrate body. Saunders, Philadelphia. 627 pp.
Russell, D.A. 1969. A new specimen of Stenonychosaurus from the Oldman
Formation (Cretaceous) of Alberta. Canadian Journal of Earth Sciences 6: 595-612.
-- 1972. Ostrich dinosaurs from the late Cretaceous of western Canada. Canadian
Journal of Earth Sciences 9: 375-402.
-- 1981. Speculations on the evolution of intelligence in multicellular organisms.
In : Life in the Universe. Edited by: J. Billingham. National Aeronautics and
Space Administration Special Publicaton, in press.
Schmidt, K.P., and R.F. Inger. 1957. Living reptiles of the world. Doubleday,
Garden City. 287 pp.
Sigmon, B.A. 1975. Functions and evolution of hominid hip and thigh musculature.
In: Primate Functional Morphology and Evolution. Edited by: R.H. Tuttle. Mouton
Publishers, The Hague. pp. 235-252.
Sokoloff, L. 1976. Circulation and energy metabolism of the brain. In: Basic
Neurochemistry , Second Edition. Edited by; G.J. Siegel, R.W. Alberts, R. Katzman,
and B.W. Arganoff. Little, Brown and Company, Boston. pp. 388-413.
Stanek, V.J. 1962. The pictoral encyclopedia of the animal kingdom. Hamlyn Publishing
Group, London. 614 pp. (Русское издание: Станек В. Я. «Иллюстрированная энциклопедия
животных» Прага, «Артия», 1983)
Sternberg, CM. 1932. Two new theropod dinosaurs from the Belly River Formation
of Alberta. Canadian Field-Naturalist 46: 99-105.
Sues, H.D. 1978. A new small theropod dinosaur from the Judith River Formation
(Campanian) of Alberta, Canada. Zoological Journal of the Linnean Society 62:
381-400.
Versluys, J. 19 23. Der Schâdel des Skelettes von Trachodon annectens
im Senckenberg-Museum. Abhandlungen von der Senckenbergischen naturforschenden
Gesellschaft 38: 1-19.
Walls, G.L. 1942. The vertebrate eye and its adaptive radiation. Cranbrook Institute
of Science Bulletin 19: 1-785.
Washburn, S.L. 1978. The evolution of man. Scientific American 239: 194-208.
Washburn, S.L., and R. Moore. 1980. Ape into human. Second Edition. Little,
Brown and Company, Boston. 194 pp.
Wellnhofer. P. 1974. Das funfte Skelettexemplar von Archaeopteryx .
Palaeontographica Abteilung A 147: 169-216.
White, T.D. 1980. Evolutionary implications of Pliocene hominid footprints.
Science 208: 175-176.
Переводчик статьи выражает глубокую благодарность Кристине Дженнес (Cristine Jennes) из Канадского музея природы за помощь в получении копий оригинальных фотографий издания и Екатерине и Андрею Сенниковым из Палеонтологического музея имени Ю. А. Орлова за помощь в переводе некоторых терминов.
Перевод: Павел Волков
г. Владимир, 2019-2020 гг.
