Одна из главных целей лабораторного занятия – проиллюстрировать тесную связь между классификацией ныне живущих видов и их эволюционными взаимоотношениями.
Классификация ныне живущих каминалькулюсов
Учащиеся начинают с того, что выстраивают 14 живых видов
в иерархическую классификацию (рис. 2). Вначале они объединяют виды в роды,
используя критерии, согласно которыми представители рода должны быть более
схожими друг с другом, чем с представителями других родов. (У каминалькулюсов,
как и у многих реальных живых организмов, физическое сходство обычно является
надёжным указанием общности происхождения.) С использованием тех же критериев
роды объединяются в семейства, семейства – в отряды, и т.д. В зависимости
от того, являются ли учащиеся «дробителями» или «объединителями» в области
таксономии, их схема классификации может остановиться на отряде, или же пройти
весь путь вплоть до типа. Если времени занятия не хватает, учащиеся могут
составить собственную предварительную классификацию дома. Ещё один способ
ускорить процесс – заставить весь класс работать над этим вопросом совместно;
при этом преподаватель выступает в роли ведущего и организатора.
Это упражнение позволяет усвоить несколько важных понятий, начиная с самого
представления об иерархической классификации. Возможно, в учебных группах
с лучшей подготовкой учителя пожелают обсудить теории классификации (Vogt
1995; Ridley 1996). Также здесь вводится понятие конвергентной эволюции, как
описано далее.
Когда учащиеся заканчивают работу по классификации, я провожу со всем классом
обсуждение с использованием проектора и изображений на прозрачной плёнке 14
ныне живущих каминалькулюсов. Я начинаю с каминалькулюса 2 и спрашиваю, какие
ещё виды принадлежат к этому же роду.
|
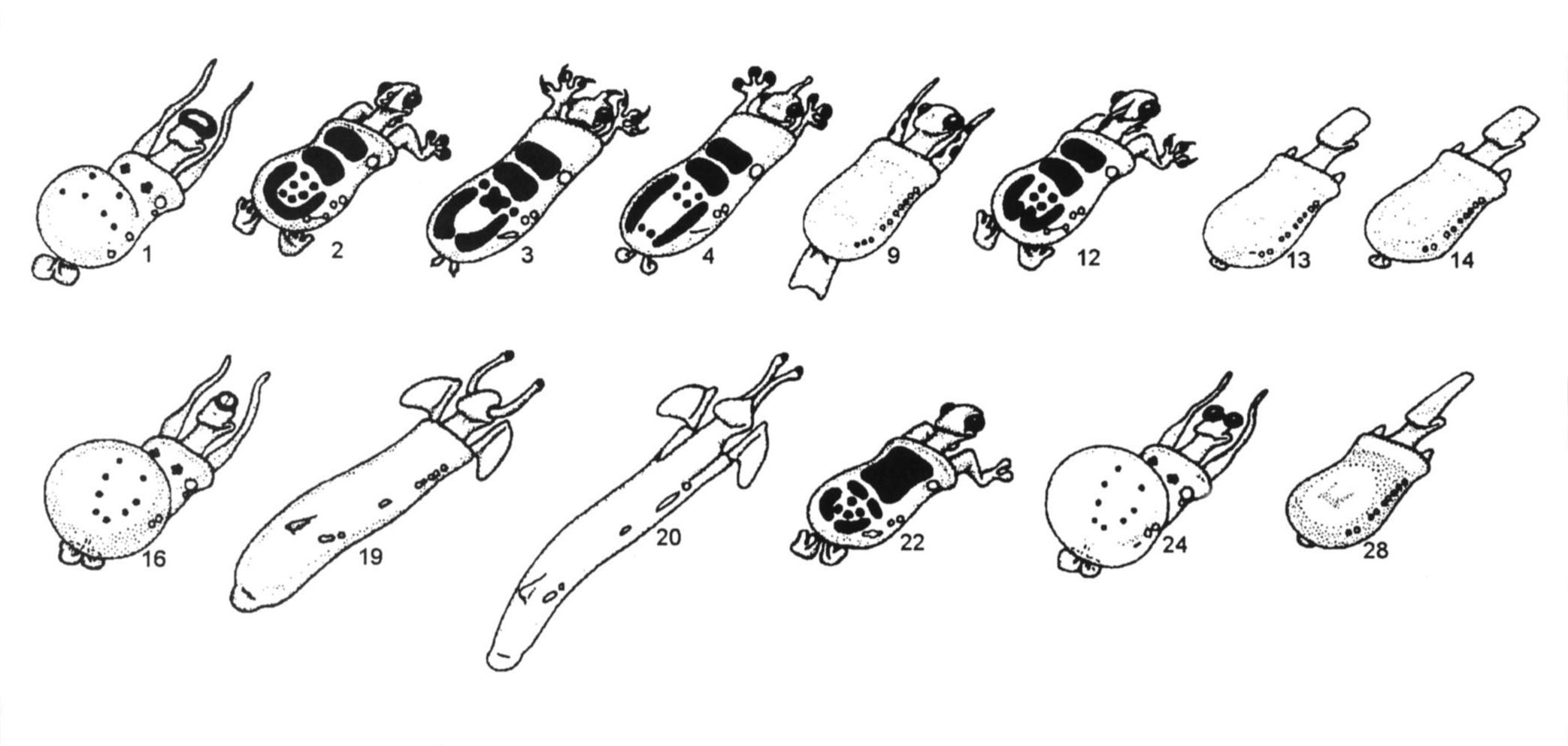
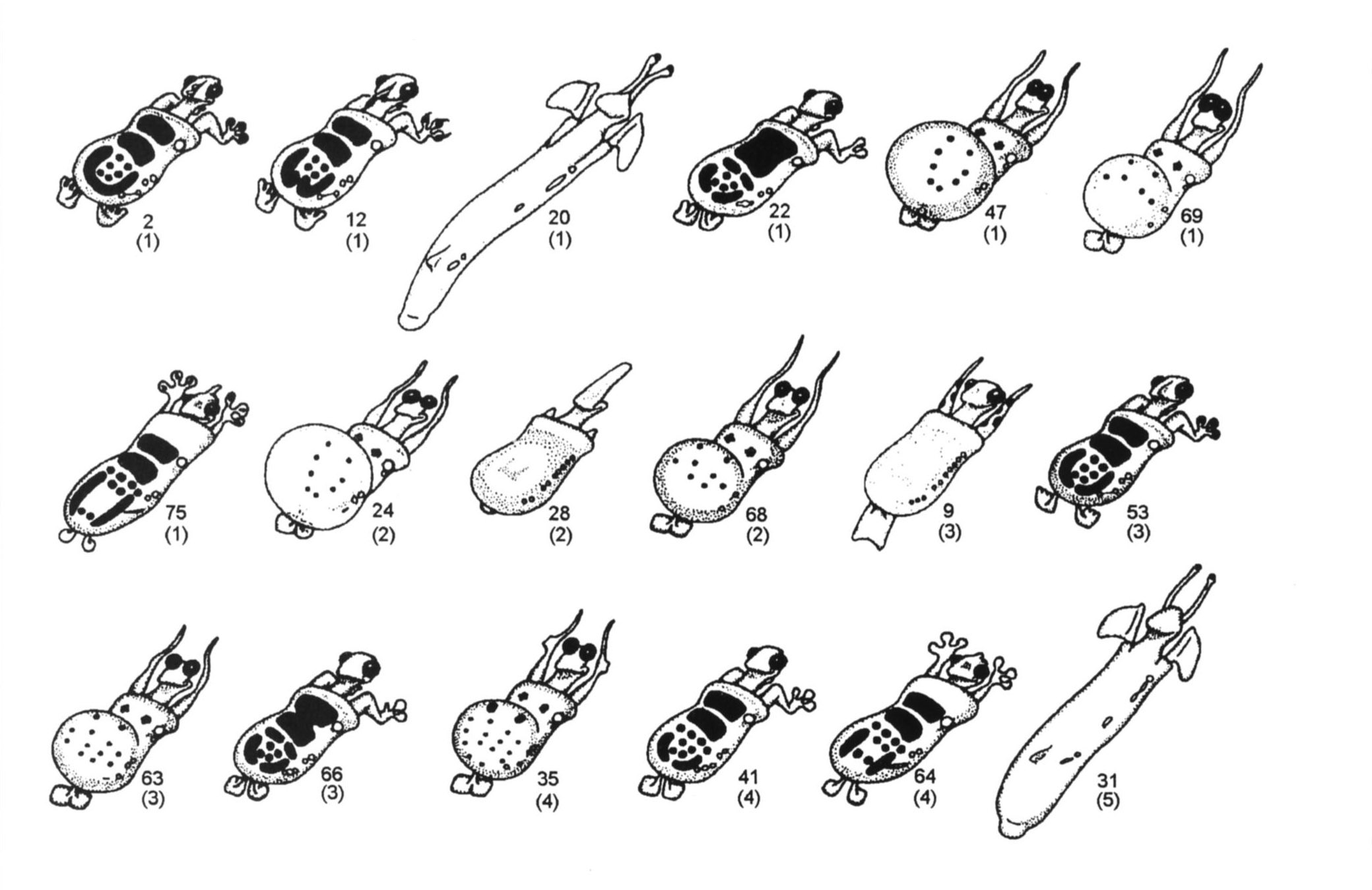
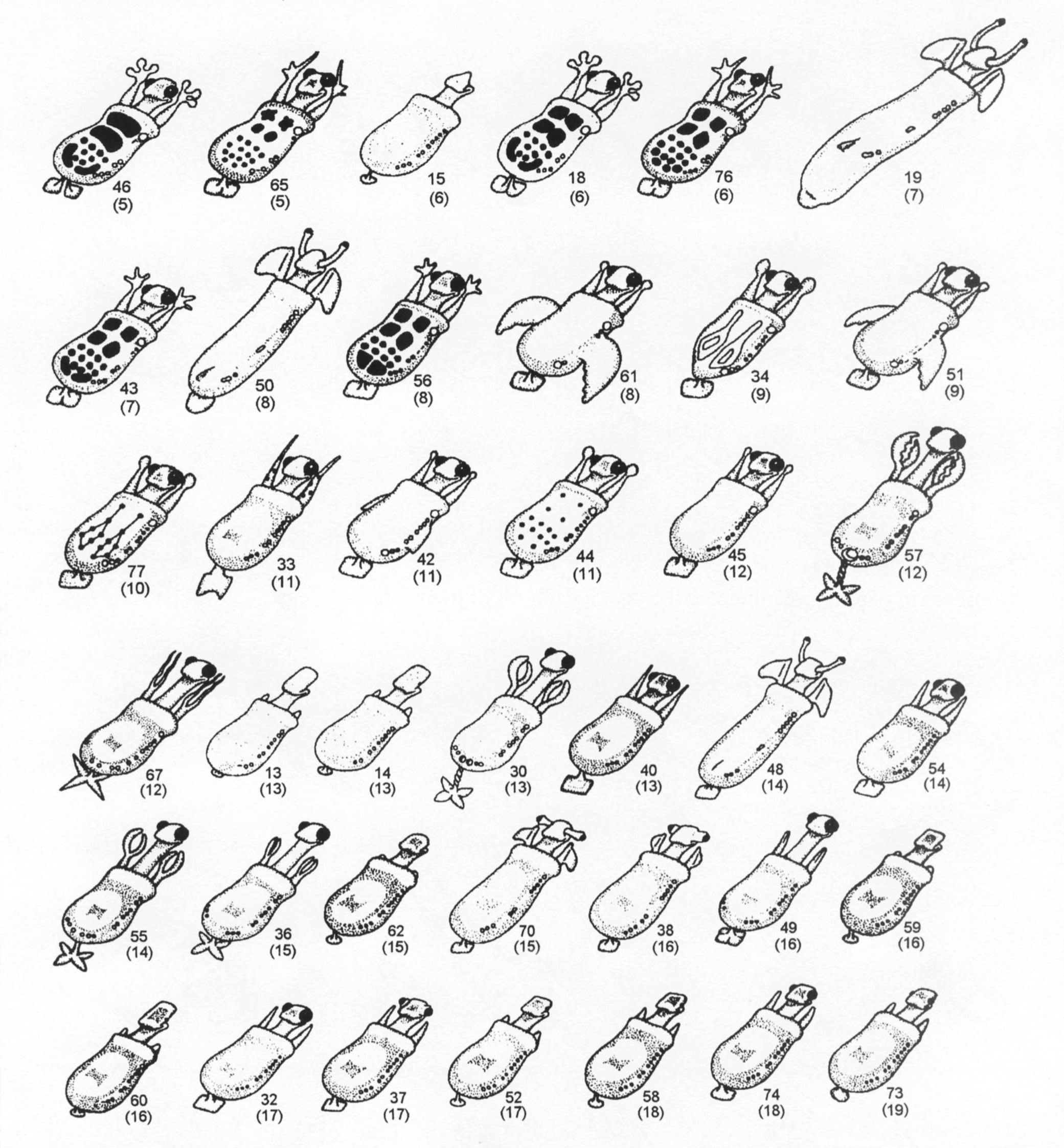
Рисунок 1. Четырнадцать ныне живущих и 57 ископаемых каминалькулюсов. Номер используется для идентификации каждого из видов вместо названия.
В большинстве своём учащиеся хотят объединить 2, 3, 4, 12
и 22 (рис. 1). Если это так, то я прошу их распределить пять видов по более
мелким родам. Самая распространённая ошибка на данный момент – это отнесение
3 и 12 в их собственный род из-за того, что у них обоих есть когти. Это даёт
возможность подчеркнуть тот момент, что классификация должна основываться
на всех доступных признаках. Когда учащиеся рассматривают такие признаки,
как цветовой узор, форма тела, наличие локтей и украшения на голове, они быстро
решают, что 3 и 4 принадлежат к одной группе (роду), а 2, 12 и 22 – к другой.
Некоторые могли бы поместить 22 в его собственный род, что вполне приемлемо.
Классифицировав пять видов ко всеобщему удовлетворению, далее мы обсуждаем
конвергентную эволюцию, используя 3 и 12 в качестве примера. (Два каминалькулюса-циклопа,
виды 1 и 16, также обладают конвергентным сходством.) Я особо отмечаю, что
о конвергентной эволюции речь идёт в том случае, если сходный признак эволюционирует
независимо в двух отдельных линиях. Например, рыбы и киты имеют одинаковую
форму, но, если отталкиваться от строения скелета, эндотермии, лактации и
т.д., киты явно принадлежат к млекопитающим. Особенности, общие для них и
для рыб, эволюционировали независимо как приспособление к жизни в водной среде.
Ещё один способ выявить конвергентную эволюцию, хотя на раннем этапе лабораторного
занятия он недоступнен для учащихся, заключается в установлении отсутствия
рассматриваемого признака у самого последнего общего предка. Это равнозначно
утверждению о том, что данные признаки являются скорее аналогичными, чем гомологичными.
Я призываю учащихся быть скорее таксономическими «дробителями», чем «объединителями»
по эвристическим соображениям; это облегчает введение понятия конвергентной
эволюции, а также подчёркивает необходимость очень тщательного изучения всех
доступных признаков. В противоположность этому, Сокал (Sokal 1983) объединяет
виды 2, 3, 4, 12 и 22 в один род.
Класс Каминалькулюсы |
||||||||||||||
Отряд |
О1 |
О2 |
||||||||||||
Семейство |
С1 |
С2 |
С3 |
С4 |
||||||||||
Род |
Р1 |
Р2 |
Р3 |
Р4 |
Р5 |
Р6 |
||||||||
Вид |
19 |
20 |
9 |
4 |
3 |
22 |
12 |
2 |
16 |
24 |
1 |
14 |
13 |
28 |
Рисунок 2. Пример иерархической классификации ныне живущих каминалькулюсов. Если взглянуть на настоящее филогенетическое древо на рисунке 2, становится очевидным, что это не лучшая схема классификации. В частности, роды 3 и 4 – это более близкие родственники рода 5, а не рода 2. Однако если не изучить сначала ископаемые свидетельства, учащиеся с гораздо большей вероятностью придут к какому-то результату вроде этого.
Филогенетическое древо, основанное только на ныне живущих видах
Во втором упражнении учащиеся используют свою классификацию
ныне живущих каминалькулюсов для построения филогенетического древа. Например,
классификация, показанная на рисунке 2, наводит на мысль о древе с рисунка
3. Одним из ключевых понятий здесь является понятие последнего общего предка.
Учащиеся узнают, что когда они относят два вида, скажем, 19 и 20, к одному
роду, это подразумевает, что у этих видов есть общий предок, которого нет
у других родов (филогенетический принцип классификации). Если род включает
три или более видов, учащиеся должны решить, какие два из этих видов обладают
самым близким родством (рис. 4).
Та же процедура применяется и на более высоких уровнях классификации. Например,
если два рода похожи друг на друга больше, чем на другие роды, это, очевидно,
связано с тем, что у них есть уникальный общий предок. Таким образом, учащиеся
узнают, что даже при отсутствии летописи окаменелостей можно разработать проект
филогенетического древа, соответствующий схеме классификации. Необязательно,
чтобы филогения точно соответствовала истинной филогении, созданной Камином,
и маловероятно, что это произойдёт, если учитывать используемый здесь интуитивный
подход. Учащиеся могут сами обнаружить какие-то ошибки после выполнения последнего
упражнения.
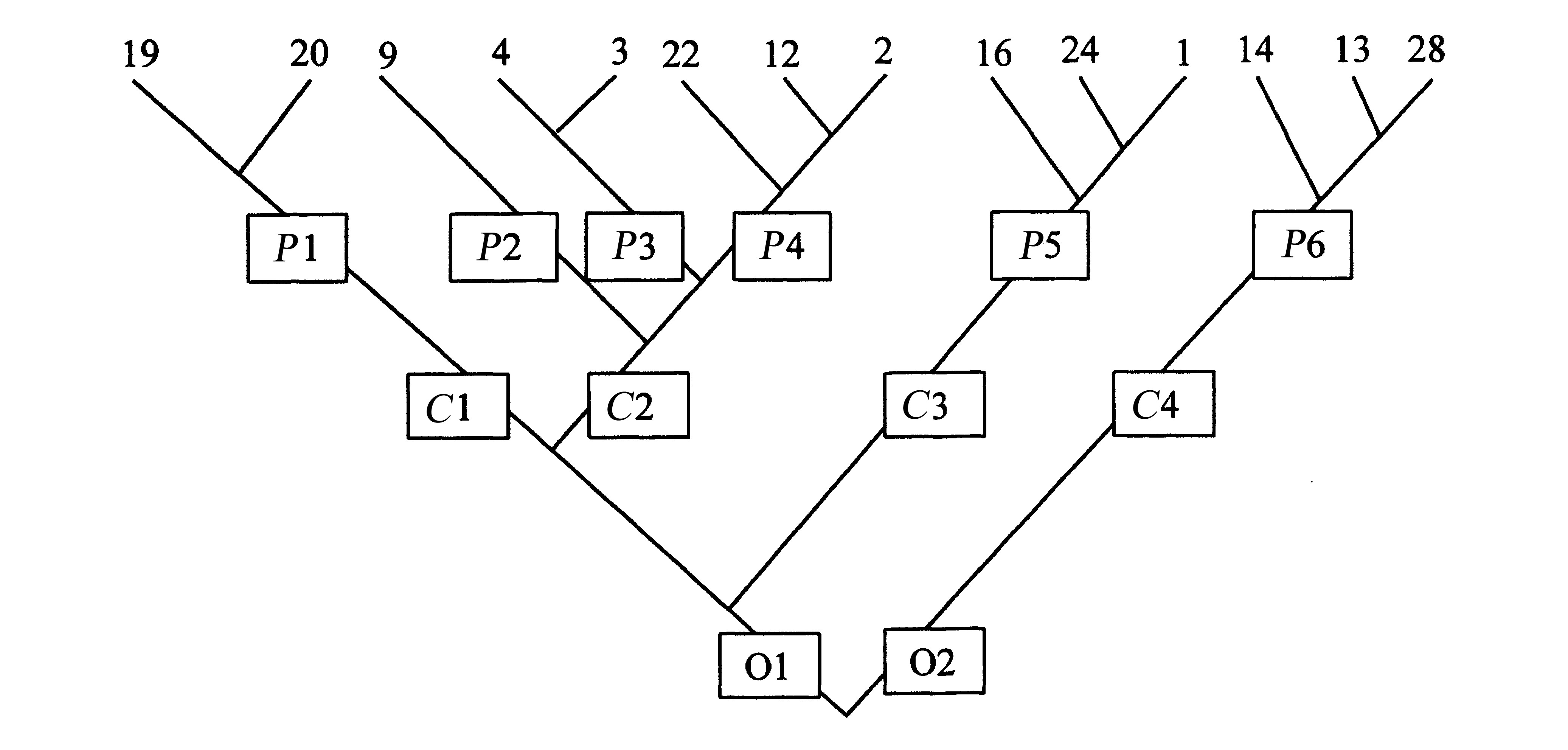 |
Рисунок 3. Это филогенетическое дерево основано на классификации ныне живущих видов, показанных на рисунке 2. У представителей каждого из родов есть общий предок, который не является общим с другими родами. То же самое верно для каждого из четырёх семейств и двух отрядов.
Филогенетическое древо, основанное на летописи окаменелостей
|
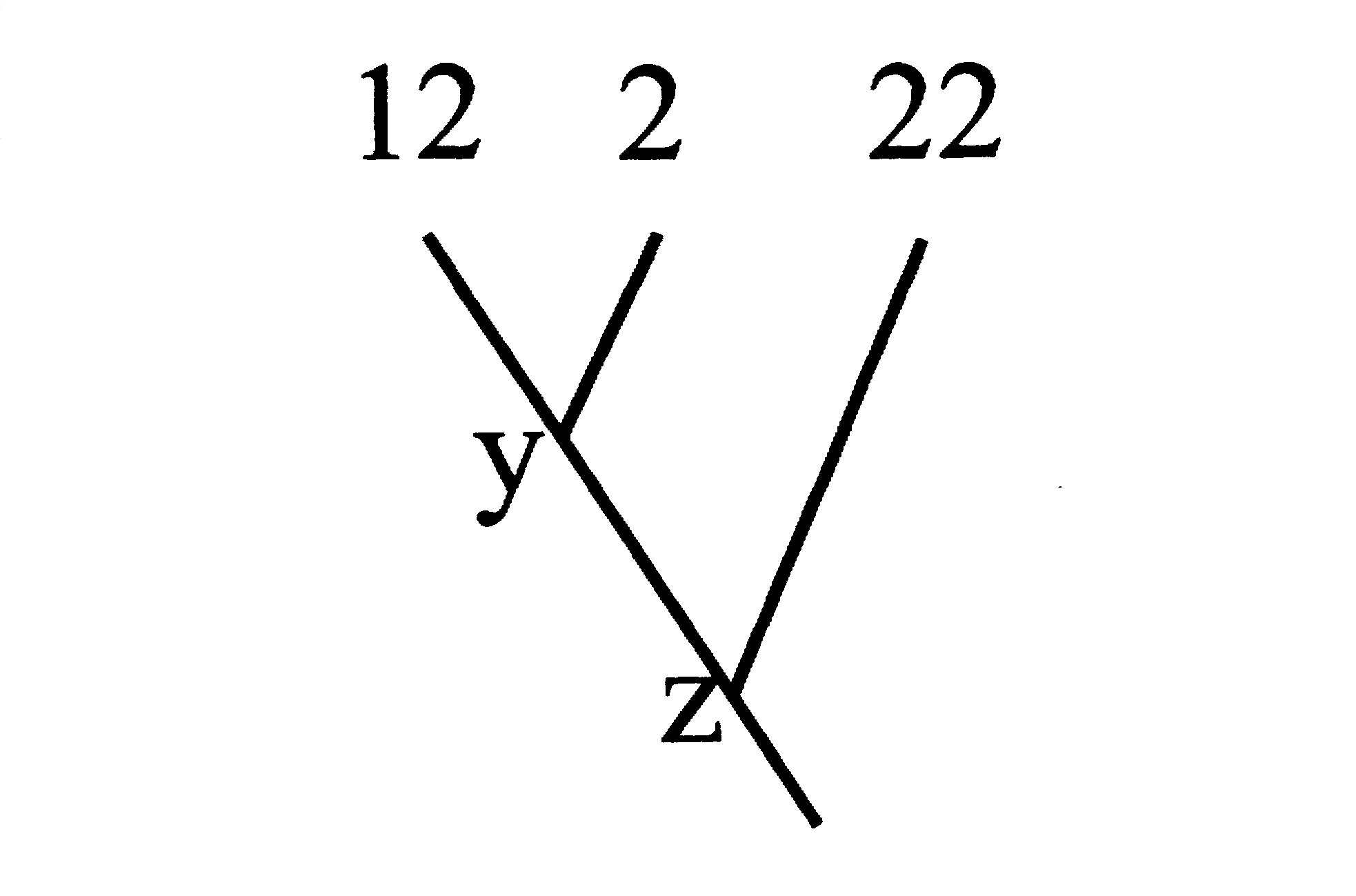
Для этого упражнения каждой группе из двух-четырёх учащихся
понадобится большой лист бумаги, ножницы для вырезания каминалькулюсов, линейка
для рисования линий на бумаге и клей для прикрепления каминалькулюсов. В качестве
бумаги мы используем остатки рулонов, которые местная газета отдаёт даром
или продает очень дёшево. Мы разрезаем бумагу на листы со стороной примерно
28 дюймов длиной.
Каждый ископаемый каминалькулюс (рис. 1) идентифицируется по номеру его вида
и возрасту (в миллионах лет) в скобках. Убедитесь, что учащиеся не отбрасывают
эти цифры. Поскольку возраст самого древнего ископаемого (вид 73) составляет
19 миллионов лет, учащиеся чертят на листе бумаги 20 горизонтальных линий
и маркируют их числами от 0 (настоящее время) вверху до 19 внизу. Обычно я
показываю классу, как начать филогенетическое древо, поместив вид 73 в середину
линии, соответствующей 19 миллионам лет. Этот вид дал начало двум новым видам
(58 и 74), представленным окаменелостями возрастом 18 миллионов лет. От этой
точки древо продолжают уже сами учащиеся. Чтобы облегчить контроль выполнения
упражнения, я использую не всех исходных каминалькулюсов. Я отсёк некоторые
из ветвей исходного древа, составленного Камином, оставив 14 ныне живущих
и 47 вымерших видов (рис. 5). Полный набор см. в Sokal (1983). Обратите внимание,
что некоторые виды представлены как живыми, так и ископаемыми образцами.
Учащимся нравится это упражнение, и они, как правило, хорошо справляются с
работой по воссозданию по кусочкам эволюции каминалькулюсов. Однако здесь
есть несколько подводных камней, с которыми могут столкнуться даже самые внимательные
из них. Эти подводные камни предоставляют прекрасную возможность обсудить
такие понятия, как пробелы в летописи окаменелостей и эволюционный застой
(рис. 5). Например, дойдя до вида 67 (возрастом 12 миллионов лет), многие
из учащихся предполагают, что он, скорее всего, произошел от вида 30 (13 миллионов
лет). Для этого был бы нужен маловероятный сценарий, когда эволюционная тенденция
к приобретению тяжёлых давящих когтей внезапно меняется на противоположную,
порождая нечто вроде раздвоенных щупальцев. Как только на этом заостряется
внимание, учащиеся сделают правильный вывод о том, что 67 ответвился дальше
внизу, и что в летописи окаменелостей существует пробел.
Завершив построение своего древа, учащиеся сравнивают его с филогенией, составленной
Камином, и устраняют какие-то обнаруженные расхождения. Затем я прошу их указать
либо в ходе лабораторной работы, либо в качестве домашнего задания:
1. Самого последнего общего предка для любых двух видов
2. Дополнительные примеры конвергентной эволюции
3. Примеры рудиментарных структур (например, внутренний палец на задней конечности
у вида 66 возрастом 3 миллиона лет)
4. Примеры эволюционного застоя (например, виды 14 и 13 остаются неизменными
на протяжении 13 миллионов лет)
5. Пример быстрой адаптивной радиации (например, множество линий, которые
происходят от вида 43 на отметке 7 миллионов лет).
Эти наблюдения можно с лёгкостью применить к примерам из реального мира, и на их основе можно сформулировать интересные вопросы для обсуждения, например:
1. Как рудиментарные структуры дают подсказки в отношении
эволюционной истории вида?
2. Какие экологические условия могут приводить к быстрому росту многообразия
в одних линиях (например, у млекопитающих в начале кайнозоя) или к длительному
застою в других (например, у мечехвостов и других «живых ископаемых»)?
3. Некоторые виды каминалькулюсов вымерли. Какие факторы могут увеличить или
уменьшить вероятность вымирания в реальном мире?
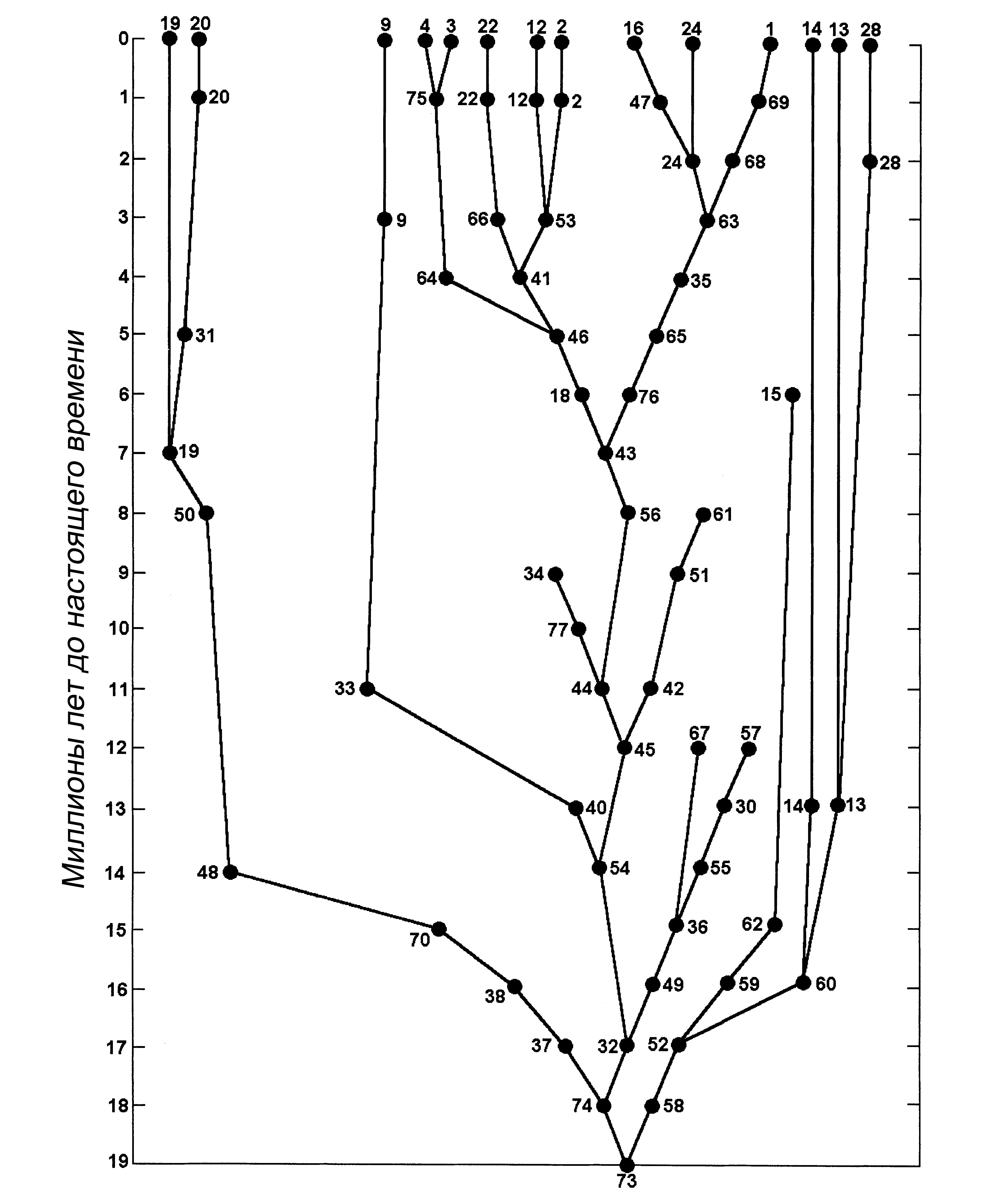 |
Рисунок 5. Эволюционное древо каминалькулюсов. Из Сокала (Sokal 1983), с изменениями. Некоторые ветви исходного древа были опущены.
Преподавателям также следует использовать древо, чтобы заострить
внимание на важном принципе: эволюционные изменения происходят путём модификации
ранее существовавших структур.
Мы использовали это лабораторное занятие в нашем курсе общей биологии для
студентов младших курсов более десяти лет (Nastase & Scharmann 1991).
Полагаю, что одна из причин его успешности заключается в том, что учащимся
нравится момент группового поиска решения проблемы в ходе выполнения упражнений.
Однако важно, чтобы инструктор отслеживал прогресс у каждой из групп, чтобы
выявлять проблемы по мере их возникновения. Это дает инструктору возможность
обсудить некоторые из понятий, упомянутых выше, с каждой из групп.
Дополнительная информация о каминалькулюсах и копия моего лабораторного задания
доступны по адресу http://www.iup.edu/-rgendron. Современные теории классификации
и филогенетического анализа более подробно рассматриваются в таких учебниках
по теории эволюции, как Ridley (1996).
Благодарности
Каминалькулы были разработаны покойным Джозефом Камином и опубликованы Робертом Сокалом. Я благодарен доктору Сокалу и редакторам журнала Systematic Biology за разрешение воспроизвести каминалькулюсов здесь и на моём веб-сайте. Я хотел бы поблагодарить многочисленных преподавателей и студентов, которые на протяжении многих лет вносили свой вклад в разработку этого лабораторного занятия. Также я благодарен B. Forbes, S. Forbes, J. McElhinney, T. Peard, S. Vogel и анонимному рецензенту за критику рукописи.